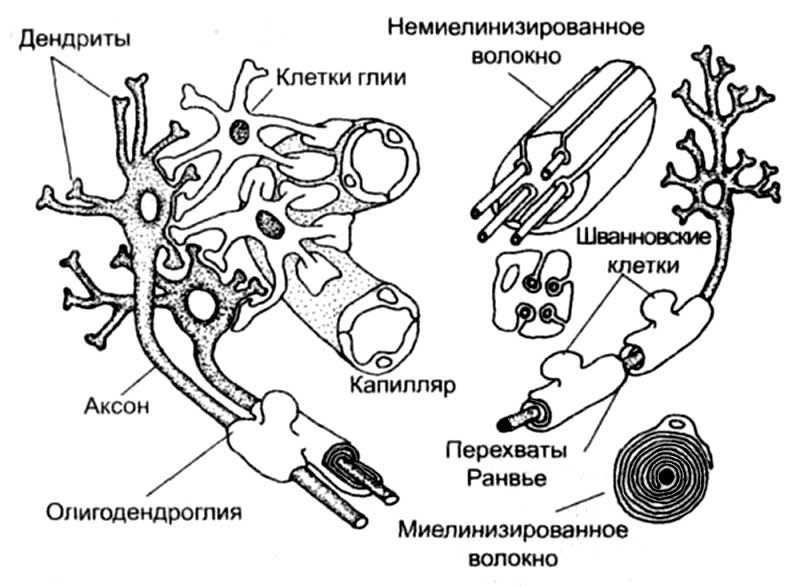
Рис. 1. Строение нейронов, клеток глии и оболочек отростков нервных клеток. В центральной нервной системе функции шванновских клеток выполняет олигодендроглия.
Нервная система — это объединение специализированных клеток, которые воспринимают, обрабатывают, хранят и используют информацию о внешней среде и внутреннем состоянии организма. Именно этим функциям подчинено строение нервных клеток. Нервные клетки имеют ряд особенностей строения, которые отличают их от других клеток организма. Нейроны неодинаковы. Они различаются по размеру, форме ветвления дендритов и аксонов, выделению различных химических веществ и физиологической активности. Однако они являются и характерными структурными элементами нервных систем всех видов многоклеточных животных. Нейроны объединяются в сети и специализированные структуры: ганглии или мозг.
У нервных клеток — нейронов — обычно можно выделить три характерные области: клеточное тело, дендриты и аксон (рис. 1). Тело содержит ядро и биохимический аппарат синтеза молекул, необходимые для жизнедеятельности клетки. Обычно тело нейрона имеет округлую, веретеновидную или пирамидальную форму. Дендриты представляют собой тонкие отростки, которые многократно ветвятся в непосредственной близости от тела клетки. Вокруг него образуется ветвистое дерево. Дендриты формируют ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксоны распространяются далеко от тела клетки. Их длина варьирует от 1 мм до 1 м, что позволяет аксонам выполнять функции линий связи между телом клетки и далеко расположенным органом-мишенью или отделом мозга. По аксону проходят сигналы, генерируемые в теле данной клетки. Аксон отличается от дендритов как по строению, так и по свойствам своей наружной мембраны. Большинство аксонов длиннее и тоньше дендритов и имеет отличный от них характер ветвления. Отростки дендритов в основном группируются вокруг клеточного тела, тогда как отростки аксонов располагаются на конце волокна, в том месте, где аксон взаимодействует с другими нейронами или органами-мишенями.
Для взаимодействия между нейронами и клетками других тканей организма у нервных клеток существуют специализированные участки мембраны, расположенные как на теле клетки, так и на ее отростках. Эти участки мембраны имеют характерное строение и называются синапсами. Через них происходят основной обмен информацией внутри нервной системы и взаимодействие нейронов с другими органами и системами организма. Нейрон может иметь от 1000 до 10 000 синапсов и получать информацию примерно от 1000 других нейронов. В типичном случае синапсы образуются между аксонами одной клетки и дендритами другой. Однако существуют и другие типы синаптических контактов: между аксоном и аксоном, аксоном и телом клетки, дендритом и дендритом, дендритом и телом клетки.
Происхождение нервных клеток в процессе эволюции животных довольно спорно. Существует две основные точки зрения на эту проблему. Наибольшее распространение получила точка зрения братьев Гертвигов (О. Hertwig, R. Hertwig, 1878—1882). Их гипотеза состоит в том, что нервные и мышечные клетки происходят из эктодермального эпителия. Они возникают независимо друг от друга и вступают во взаимодействия уже вторично (рис. 2,а—в). В соответствии с гипотезой Гертвигов источником всех нервных клеток является первичная чувствительная клетка, возникшая из эктодермы и получившая возможность воспринимать раздражения и генерировать и проводить возбуждение. Специализированные отростки этих клеток вступают в связь с независимо возникшими мышечными клетками. По мнению авторов этой гипотезы, мышечные волокна возникли из клеток, сходных с эпителиальномышечными клетками кишечнополостных. Такой простейший нервно-мышечный комплекс развивается по пути дифференциации нервного аппарата, что приводит к возникновению вставочных нейронов или ганглиев (см. рис. 2,в). Нервная система подразделяется на чувствительные клетки, лежащие в эктодерме, и вставочные нейроны, обладающие интегративной функцией. Последние являются предвестниками централизации обработки сенсорной информации и возникновения мозга.

а-в — гипотеза братьев Гертвигов;
г-е — гипотеза Клейненберга-Заварзина.
По гипотезе Гертвигов нервные и мышечные клетки происходят независимо из эктодермы, а затем объединяются в нервно-мышечный комплекс.
По представлениям Клейненберга-Заварзина из эктодермы возникает клетка, обладающая сенсорными и моторными функциями одновременно. Затем она дифференцируется на две системы: нервную и мышечную.
Штриховка — мышечные клетки или волокна, точки — чувствительные или нервные клетки.
Другая точка зрения на происхождение нервных клеток сформировалась в работах Клейненберга и Заварзина (Н. Kleinenberg, 1872; А.А. Заварзин, 1941). Ее суть сводится к тому, что нервная и мышечная системы рассматриваются как единое и нераздельное целое. Считается, что обе системы возникли одновременно в виде такого образования, которое не имело дифференцированных мышечных и нервных компонентов С точки зрения этой гипотезы первичными были клетки, выделившиеся из эктодермы и обладавшие как нервными, так и мышечными функциями (рис. 2,г,д). Часть клетки обладала способностью к чувствительности, а другая — к сокращению. В гипотезе подчеркивается первичность морфофункциональной связи между нервными и мышечными клетками. В дальнейшем, по мнению авторов, произошла дифференциация чувствительно-сократимых клеток на чисто сенсорные и моторные, а общность их происхождения стала основой нервно-мышечных взаимодействий (рис. 2,е).
Нейроны — это морфологические единицы, кирпичики, из которых построена нервная система как позвоночных, так и беспозвоночных животных. Сходство строения и функций нейронов объединяет все группы организмов, обладающих нервной системой. Нейрон является минимальной единицей в анализе нейробиологических процессов. Ионы и молекулы, находящиеся в клетке, равно как и органеллы, обеспечивающие функционирование нейронов, сходны с аналогичными структурами в других органах и тканях организма животных и человека. Их не стоит принимать за низший уровень организации нервной системы, как нельзя учитывать открывание и закрывание дверей купе пассажирского поезда при составлении расписания поездов на конкретном участке железной дороги. Как поезд является единицей в расписании движения поездов, так и нейрон представляет собой минимальную единицу в организации нервной системы. Нервные клетки объединены в нервные системы различным образом. В простейшем случае эти элементы распределены вполне равномерно по всему телу животного или по большей его части. Это диффузное распределение нервных клеток. Являясь наиболее простым, оно в то же время оказывается филогенетически наиболее старым. В этом случае клетки расположены однородно и снабжены многочисленными одинаковыми отростками, которые объединены в общую сеть (рис. 3,а). Протяженных и организованных путей в такой сети нет, отростки клеток связывают только соседние нейроны. В сеть входят волокна, идущие от воспринимающих клеток эпителия или рецепторов, а из сети отходят двигательные волокна, оканчивающиеся на поверхности мышечных клеток.
Другой вариант представляет собой компактную организацию нервной системы. В этом случае клеточные тела передаточных и двигательных нейронов собраны в небольших зонах тела, которые называются нервными ганглиями или узлами. Для беспозвоночных характерны скопления нервных клеток, не содержащие внутреннюю полость (см. рис. 3,а,б). Внутренняя зона ганглия состоит из нервных клеток, окруженных нейропилем, представляющим собой сплетение отростков дендритов этих клеток. В ганглиях расположены чувствительные нейроны или оканчиваются их аксоны, что позволяет ганглиозным клеткам получать информацию с периферии тела беспозвночного. Аксоны двигательных нейронов, выходящие из ганглиев, образуют более или менее длинные проводящие пути, связывающие с нервными центрами органы дыхания, пищеварения, железы и мышечные клетки.
Для хордовых характерно появление более сложной структуры организации нервной системы. Наряду с сетями и ганглиями у позвоночных формируется центральная трубчатая нервная система. Возникает головной и спинной мозг с полостью внутри, которая называется спинномозговым каналом или мозговыми желудочками (рис. 3,в). Размеры центральной трубчатой нервной системы позвоночных позволяют разместить между чувствительным и двигательным нейроном большое количество вставочных нейронов, которые обрабатывают получаемую информацию. От качества переработки этой информации зависит сложность поведения животного или интеллектуальная деятельность человека.
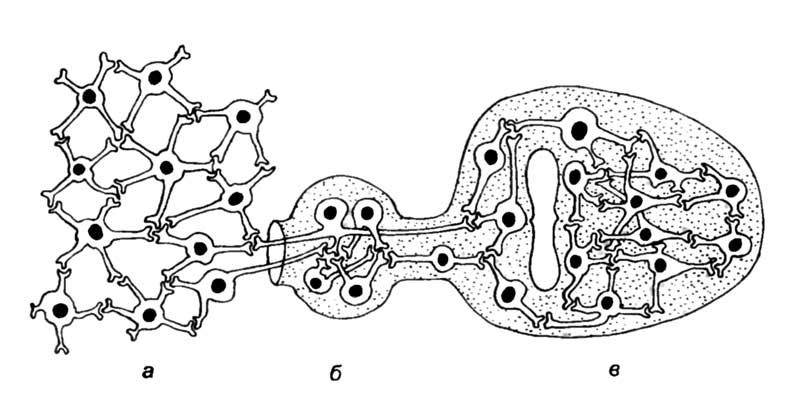
а — диффузный тип строения, характерный для кишечнополостных;
б — ганглиозный тип организации нервной системы, который характерен для червеобразных, моллюсков и членистоногих;
в — центральная трубчатая нервная система хордовых, в которой присутствуют все типы интеграции нейронов.
Эти общие принципы организации нервной системы характерны как для беспозвоночных, так и для позвоночных животных. Однако существует ряд принципиальных различий, которые следует подчеркнуть.
Во-первых, строение нейронов нервной системы беспозвоночных и позвоночных существенно различается как по размерам, так и по форме (рис. 4).
Во-вторых, для большинства беспозвоночных характерно отсутствие полости внутри мозга — мозговых желудочков. Исключением являются головоногие моллюски, головные ганглии которых формируют внутренние полости.
В-третьих, у беспозвоночных стратификация нейронов распространена значительно менее, чем у позвоночных.
Эти морфологические различия приводят к тому, что эволюция нервных систем позвоночных и беспозвоночных животных происходила в рамках очень несхожих ограничений. Для беспозвоночных с их маленькой нервной системой было бы невозможным доминирование регуляционного эмбрионального развития нервной системы над генетически детерминированным. Это связано с тем, что при регуляционном развитии судьба эмбриональной клетки вероятностна и зависит от межклеточных взаимодействий. В таком случае требуются некоторый переизбыток нейробластов, их конкурентное поведение при дифференцировке и программированная гибель клеток. Такой переизбыток эмбрионального материала практически невозможен у беспозвоночных. В результате возникла ситуация, когда нервная система развивается преимущественно по детерминационному типу. Это означает, что число нервных клеток, их связи и даже ветвление дендритов предопределены генетически. Если попытаться выразить соотношение детерминированных и регуляционных событий в развитии нервной системы беспозвоночных количественно, то оно составит примерно 7:3. Понятно, что при таком соотношении практически не остается места для морфологической индивидуализации нервной системы. Более того, достаточно жесткая детерминированность организации нервной системы создает возможность эффективной передачи по наследству разнообразных форм поведения. Поэтому мы встречаем среди беспозвоночных преимущественно инстинктивное поведение, его незначительную индивидуализацию и отсутствие «высоких» психологических свойств. Не менее важным морфологическим и функциональным отличием беспозвоночных является система нейро-гормональной регуляции поведения. Нервная и эндокринная системы образуют интегрированные нейроэндокринные комплексы. У наиболее эволюционно «продвинутых» видов беспозвоночных в рефлекторные реакции и обработку сенсорной информации входит нейрогормональный этап. Удельный вес нейрогормонального этапа в работе головных ганглиев может различаться, но его значение у беспозвоночных несопоставимо выше, чем у позвоночных животных. Для большинства высших беспозвоночных характерно формирование специального нейрогормонального органа, расположенного позади головных ганглиев. Эти образования носят название нейрогемального органа, а при интеграции с головными ганглиями или нервами — нейрогемальной зоны. Наличие этого центра приводит к тому, что поведение животного при любой активизации головных ганглиев начинает контролироваться выделяемыми гормонами. Следовательно, оно становится более генерализованным, программируемым и предсказуемым. Соотношение между влиянием нейрогормональных и нервных комплексов на поведение специально не изучали. Однако приблизительная оценка показывает, что у высших беспозвоночных поведение примерно на 85% контролируется нейрогормонами, тогда как у позвоночных доля этого влияния не достигает 50%.
Таким образом, сопоставляя историческое развитие нервной системы, необходимо отметить, что наибольшего результата эволюция достигла у беспозвоночных животных. Их нервная система очень небольшого размера, обладает развитыми сенсорными входами и имеет практически полный набор программ поведения, находящийся под контролем генетических или нейрогормональных процессов. Эта компактность, экономичность и детерминированность делают нервную систему беспозвоночных совершенным инструментом для решения стандартных задач и коллективных действий. Учитывая однотипность и предопределенность реакций нервной системы, легко понять согласованность миграций, половую и суточную активности высших беспозвоночных. Совершенство нервной системы беспозвоночных позволило им освоить огромные пространства и быть самой распространенной группой животных на планете.
Однако существует и обратная сторона медали. За компактность, экономичность и эффективность нервной системы беспозвоночным пришлось заплатить индивидуализацией поведения. Беспозвоночные практически не обладают внутривидовой изменчивостью строения нервной системы и как следствие индивидуальными особенностями поведения. Они совершенны в предусмотренных природой случаях и беспомощны в нестандартной ситуации. Из высших беспозвоночных получились бы идеальные солдаты, но не появился бы ни один генерал. Там, где начинаются индивидуальность и творчество, царят морфологическая изменчивость и случайность.
Нервная система позвоночных построена на принципах вероятностного развития, дублирования, избыточности и индивидуальной изменчивости. Это не означает, что в мозге позвоночных нет места генетической детерминации развития или нейрогормональной регуляции. Все эти слагаемые поведения присутствуют и в организации нервной системы позвоночных, однако они играют несоизмеримо меньшую роль, чем у беспозвоночных. При развитии нервной системы позвоночных действуют законы формообразования, которые детерминируют первичную региональную экспрессию генов и морфогенез (С.В. Савельев, 1993). Одновременно присутствует и огромная переизбыточность эмбриональных нейробластов. Строгой детерминации развития каждого отдельного нейрона, как у беспозвоночных, нет. Судьба каждой клетки носит вероятностный характер и зависит от тех коммуникативных взаимодействий, в которые она вступит во время своего индивидуального развития и дифференцировки. Достаточно высокая случайность судьбы каждого нейробласта отражается в огромных масштабах гибели клеток нервной системы. При развитии головного мозга млекопитающих нормальная гибель клеток обычно составляет 0,2—0,4%. К концу метаморфоза бесхвостых амфибий гибель клеток достигает 85% от общего числа нейробластов спинного мозга. Такие потери клеток для беспозвоночных были бы невосполнимы, тогда как у позвоночных эти события составляют нормальный морфогенез. Вероятностное развитие нервных клеток позвоночных подтверждено в многочисленных экспериментах (С.В. Савельев, 1993). Достаточно упомянуть способность нервной системы позвоночных к компенсации искусственно вызванной гибели клеток. Если у эмбриона амфибии на стадии нервной пластинки случайным образом (не в одном месте) разрушить 40% клеток, то зародыш разовьется в нормальный организм немного меньшего размера, чем обычная особь. Следовательно, нервная система позвоночных уже в эмбриональный период закладывается с «переизбытком» клеток, поскольку их судьба не столь строго детерминирована, как у беспозвоночных. Таким образом, известные различия в развитии нервной системы позвоночных и беспозвоночных животных позволяют назвать онтогенетическое развитие позвоночных регуляционным, а беспозвоночных — детерминационным. Это не означает полного отсутствия детерминационных явлений у позвоночных и регуляционных процессов у беспозвоночных. Однако явное превалирование одного процесса над другим совершенно очевидно.
Различные стратегии развития нервной системы позвоночных и беспозвоночных отражаются и на основных принципах ее морфологической организации. Если нервная система компактизована из-за размеров и массы беспозвоночных, то у позвоночных нет физических препятствий для ее увеличения. Отсутствие ограничения на размер нервной системы позволяет достигать головному и спинному мозгу позвоночных крупных размеров и массы до 10 кг. Однако различия в размерах мозга не исключают сходство гистологической организации между позвоночными и беспозвоночными.
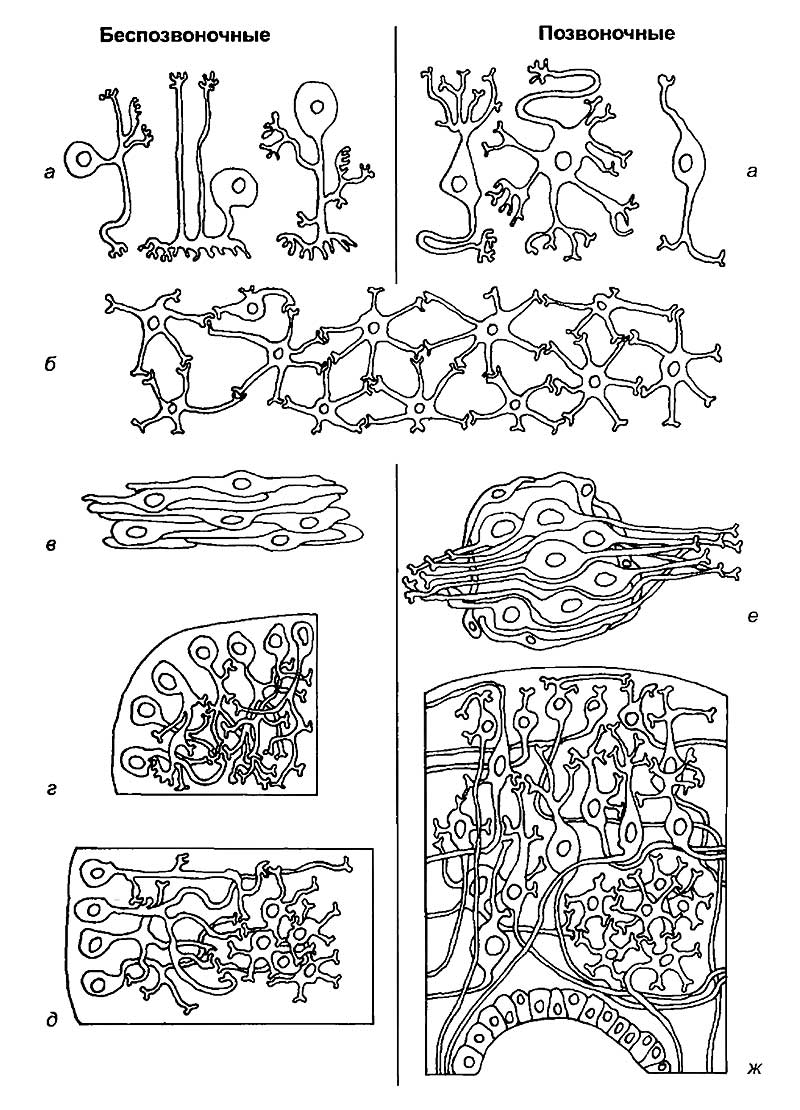
а — типы нейронов;
б — диффузная сеть, характерная для обеих групп;
в — «параллельные ганглии»;
г — корково-нейропильный тип организации;
д — островковая или ядерная форма концентрации нервных клеток в нейропиле;
е — периферический ганглий позвоночного;
ж — кора, подкорковые ядра и стенка мозговых желудочков.
Нервные клетки в обеих группах не имеют принципиальных различий в цитологической организации (см. рис. 4), хотя некоторые особенности в строении отростков и тел клеток могут быть предметом исследования сравнительной цитологии. В самом простом случае нервные клетки образуют диффузную сеть, характерную для обеих групп (см. рис. 4,б). В ганглиях беспозвоночных и нервной трубке позвоночных клетки и их отростки расположены закономерным образом. К первому типу организации нервных клеток беспозвоночных следует отнести их способность формировать «параллельные ганглии» (см. рис. 4,в). Эти структуры характеризуются однонаправленным вытягиванием тел нейронов и формированием контактов между параллельно лежащими отростками.
Второй тип организации нейронов наиболее характерен для беспозвоночных и носит название дифференцированного нейропиля (см. рис. 4,г). Он состоит из тел клеток, расположенных в корковом слое, и нейропиля, состоящего из переплетенных отростков нейронов. Это наиболее распространенный принцип строения туловищных и головных ганглиев.
Третьим типом концентрации нервных элементов у беспозвоночных является нейропиль с островками тел клеток (см. рис. 4,д). Локальные концентрации тел клеток беспозвоночных сходны с ядрами головного и спинного мозга позвоночных. Они характеризуются компактным расположением тел нейронов внутри нейропиля, что позволяет клеткам эффективнее обмениваться информацией между собой.
Для позвоночных характерно наличие периферических ганглиев, окруженных соединительнотканной оболочкой (см. рис. 4,е). Тела клеток в таких ганглиях расположены по всему объему структуры и не имеют поверхностной локализации, как у беспозвоночных. Нейропиль, состоящий из переплетенных отростков клеток, не выражен. Корковые структуры позвоночных характеризуются многослойным расположением тел клеток (см. рис. 4,ж). Они обычно разделены пучками волокон, которые позволяют выделять как горизонтальные слои коры, так и вертикальные колонки. Для позвоночных специфично участие разнообразных типов клеток в формировании коры, тогда как для беспозвоночных это довольно редкое явление. Принципиальным отличием этих групп является наличие мозговых желудочков у позвоночных, которые окружены специализированными эпендимными клетками. Эти клетки сохраняют свои эмбриональные свойства и могут участвовать в регенерации мозга. У постоянно растущих животных, таких, как неотенические виды хвостатых амфибий, акулы и некоторые рептилии, эпендимные клетки спинного мозга могут пролиферировать и формировать отростки у половозрелых животных. Для некоторых певчих птиц характерны сезонная пролиферация эпендимных клеток и их миграция в «певческие» центры мозга самцов. Таким образом происходит формирование морфологического субстрата для усложнения песенного репертуара. Сохранение эпендимными клетками способности к пролиферации является подтверждением регуляционного принципа организации мозга позвоночных. Подобная «резервная избыточность» для беспозвоночных неизвестна.
Эти данные свидетельствуют не только о глубоких различиях, но и о сходстве развития и организации нервной системы позвоночных и беспозвоночных. Реконструируя пути усложнения морфологической организации мезга, можно предположить следующую последовательность событий. На первом этапе исторического развития нервной системы произошло появление чувствительных элементов из клеток эктодермального зачатка (рис. 5,а,б). Специализированные клетки эктодермы одновременно обладали сенсорными и эффекторными функциями. Они рецептировали сигнал, проводили его к эффектор-ным органам и запускали их реакцию. Эти клетки были связаны между собой и формировали непрерывную сеть, которая не имела выраженных центров (рис. 5,в). Такой тип организации нервной системы мы встречаем у кишечнополостных. При появлении более сложных поведенческих задач элементы нервной системы стали объединяться в небольшие скопления. По-видимому, это происходило двумя путями. С одной стороны, формировались «параллельные ганглии» (рис. 5,г), для которых характерны синаптические контакты между телами клеток. Этот примитивный тип концентрации нервных элементов характерен для свободноплавающих кишечнополостных.
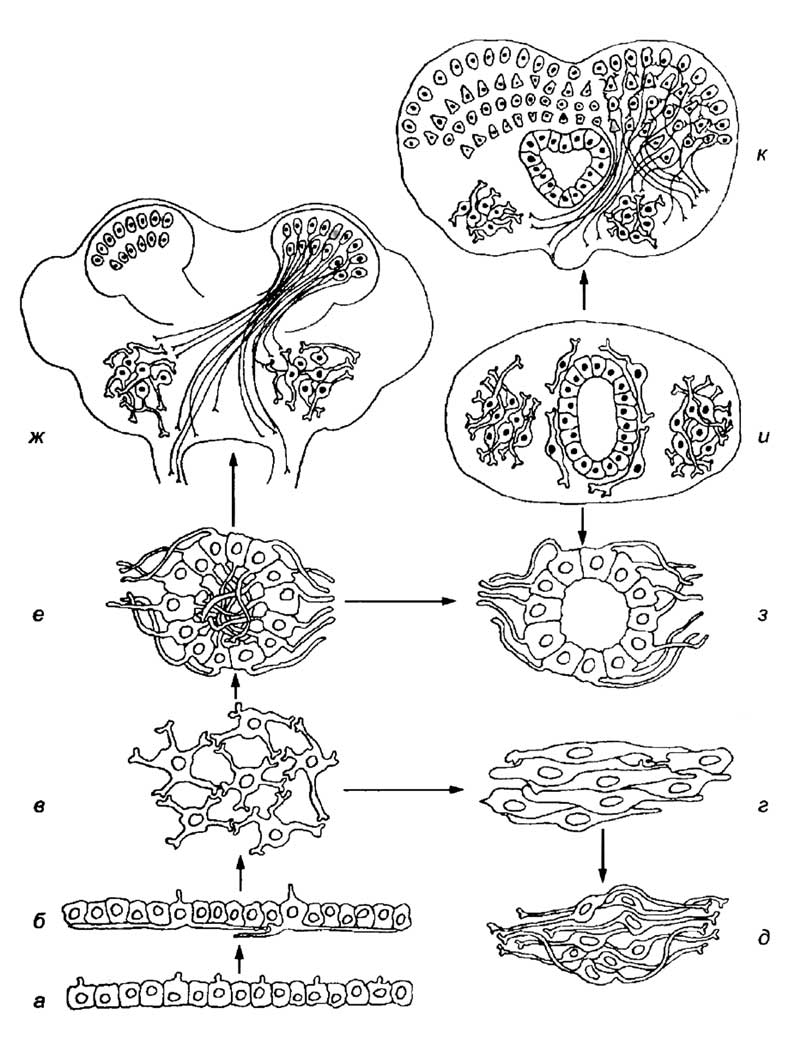
а — эктодерма без специализированных чувствительных элементов;
б — эктодерма, содержащая нервные клетки, связанные между собой отростками;
в — диффузная нервная сеть, не содержащая выраженных центров;
г — «параллельные ганглии» беспозвоночных;
д — ганглии без выраженного нейропиля;
е — ганглии с нейропилем;
ж — головной ганглий беспозвоночного с грибовидными телами, которые являются стратифицированными структурами и островковыми (ядерными) скоплениями тел нейронов;
з — нервная трубка, образовавшаяся за счет выхода отростков нейронов на наружную поверхность нервого тяжа;
и — нервная трубка с ядрами и прижелудочковым расположением нейронов;
к — стратифицированные структуры позвоночных коркового типа.
С другой стороны, появились скопления нейронов с наружным расположением тел клеток и нейропилем из переплетенных отростков внутри ганглия (рис. 5,е). Этот тип организации ганглиев оказался достаточно эффективным и сохранился до настоящего времени у большинства беспозвоночных. Такой ганглий обладает рядом преимуществ, которые имеют особое значение для животных с незамкнутой кровеносной системой. Тела его нейронов расположены преимущественно на наружной поверхности, что позволяет поддерживать довольно высокий уровень метаболизма: снабжать питательными веществами, кислородом и отводить токсичные продукты экскреции клеток. Ней-ропиль, находящийся внутри ганглия, дает возможность обмениваться сигналами и формировать генерализованный ответ на разнообразные сенсорные сигналы. По-видимому, из этой формы концентрации нервных элементов возникли головные и туловищные ганглии высших беспозвоночных (рис. 5,ж), ганглии и нервная трубка позвоночных (рис. 5,д,з). В головных ганглиях беспозвоночных сложились два основных типа гистологических структур: островковые скопления тел клеток и стратифицированные грибовидные тела. Островковые скопления тел клеток беспозвоночных практически идентичны подкорковым и стволовым ядрам позвоночных. Организация грибовидных тел напоминает слоистое расположение клеток в коре млекопитающих. Однако грибовидные тела беспозвоночных не имеют упорядоченных вертикальных связей между нейронами и колончатой организации. Тем не менее стратификация нейронов в грибовидных телах предполагает сходство механизмов обработки информации в ассоциативных центрах позвоночных и беспозвоночных животных.
По-видимому, нервная система позвоночных возникла из ганглиев беспозвоночных с нейропилем из переплетенных отростков (см. рис. 5,е). Трубчатая нервная система сформировалась в результате выхода отростков нейронов из внутренней полости ганглия (см. рис. 5,е,з). Это событие привело к формированию нейропиля из отростков нервных клеток с наружной стороны нервной трубки. На следующем этапе формирование новых нервных центров происходило преимущественно вокруг желудочков. Это привело к возникновению центрального серого вещества. Часть клеток выселялась из прижелудочковой зоны и формировала структуры ядерного или стратифицированного типа во внешнем нейропиле (рис. 5,и,к). У высших позвоночных центральное серое вещество практически отсутствует, а основные нервные центры мозга представлены сложными ядрами и корковыми структурами различных типов. В дальнейшем цефализация позвоночных развивалась по принципу количественного наращивания анатомического представительства анализаторных систем и ассоциативных центров. В отличие от беспозвоночных нервная система трубчатого типа при наличии замкнутой кровеносной системы может бесконечно увеличиваться в размерах. Это позволило позвоночным достигнуть крайне высокого уровня развития умственных способностей.
Подводя итог краткому обзору основных принципов анатомической и гистологической интеграции нервных клеток у беспозвоночных и позвоночных животных, необходимо сделать несколько общих выводов. Во-первых, нейроны в обеих группах имеют сходное строение, но различаются по размерам. Во-вторых, интегративные взаимодействия между нейронами осуществляются в сходных гистологических образованиях: ганглиях, ядрах и стратифицированных структурах. Эти образования встречаются как у беспозвоночных, так и у позвоночных животных. Основными морфологическими отличиями позвоночных являются система мозговых желудочков и организация кровеносной системы. Эффективная система кровоснабжения позволяет поддерживать высокий уровень метаболизма нервной системы. Обширная система мозговых желудочков и сосудистое сплетение обеспечивают осмотический и водный баланс головного и спинного мозга. Эти морфологические особенности позвоночных обеспечивают лабильность, высокие адаптационные возможности и большие размеры нервной системы. Нервная система беспозвоночных более консервативна, имеет небольшие размеры и не обладает столь широкими адаптационными возможностями, как у позвоночных. Зато она имеет огромный набор генетически детерминированных программ поведения, четко запрограммированное развитие и эффективную систему гормональной регуляции физиологической активности. При небольших размерах беспозвоночных это позволяет добиваться впечатляющих успехов в конкуренции с другими видами.
Таким образом, сравнение организации позвоночных и беспозвоночных отражает различные принципы развития нервной системы, что дает представление о возможных вариантах ее строения и путях эволюции.
Основной особенностью строения кишечнополостных является радиальная симметрия. Форма тела кишечнополостных в основном напоминает чашу, иногда уплощенную или удлиненную с простой полостью внутри, сообщающейся с внешней средой единственным отверстием, одновременно выполняющим функции рта и ануса. Органы у кишечнополостных отсутствуют, что говорит о тканевом уровне организации этих животных. У них существует только два клеточных слоя со студенистой мезоглеей между ними (рис. 6).
Почти у всех кишечнополостных нервная система представлена диффузной сетью нейронов с отростками, лишенными оболочки. В наиболее чистом виде мы встречаем ее у прикрепленных форм (см. рис. 6). У всех кишечнополостных нервная сеть залегает в слое неклеточного вещества — мезоглее, расположенном между двумя слоями клеток, составляющих стенку тела. Из наружного слоя — эктодермы — нервная сеть получает информацию от хеморецепторных сенсорных ямок, светочувствительных глазков, сенсорных тактильных клеток и гравитационных рецепторов — статоцитов. Сигналы от этих рецепторов распространяются по нервной сети и идут к сократительным эпителиальным, железистым и пищеварительным клеткам гастродермы — внутреннего слоя. У этого общего для кишечнополостных типа организации существуют многочисленные отклонения.
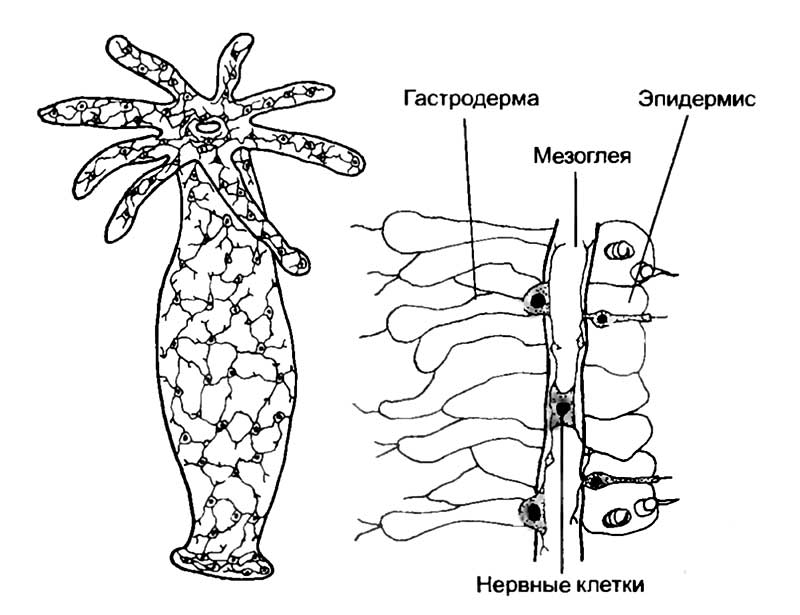
Для гидры характерна диффузная нервная система с плотным расположением нейронов на подошве и у ротового отверстия. Увеличен фрагмент стенки тела гидры с закрашенными нервными клетками, которые расположены в мезоглее и на границе гастродермы и эпидермиса.
У актиний нервная сеть не везде одинакова. Нервные клетки встречаются чаще у оснований щупалец и по краю ротовой площадки. Еще более неоднородна нервная система у свободноплавающих кишечнополостных. Нервная сеть медуз ограничена нижней поверхностью зонтика и желудочного стебелька. У гидромедуз по краю зонтика формируется двойное нервное кольцо, соединенное нервами с органами равновесия, хеморецепторами и сенсорными тактильными клетками. Централизация нервной системы особенно заметна у сцифоидных медуз. Их нервное кольцо состоит из многочисленных нервных клеток, сконцентрированных в 8 краевых тельцах. Эти данные свидетельствуют о том, что уже на уровне кишечнополостных происходит концентрация нервных клеток в ганглиоподобные структуры.
Различия в организации нервной системы реализуются в поведении кишечнополостных. При сильном раздражении гидры или актинии происходит быстрое сокращение всего тела животного. Это общая генерализованная реакция диффузной нервной системы. Интересно отметить, что если у актинии вырезать участок тела или отделить щупальце, то на протяжении суток оно будет продолжать реагировать на раздражения, как и в теле животного. Следовательно, диффузная нервная система представляет собой однородное образование, не имеющее более или менее важных зон или центров. Даже такая просто устроенная нервная система может быть причиной весьма целесообразного поведения животного. Говоря о гидромедузах, необходимо отметить, что их нервная система уже отличается от исходного диффузного типа. Кольцевые нервные тяжи, проходящие по краю зонтика, одновременно включают несколько нейронов и их отростков. Рассеянные тела нейронов имеют отростки, вытянутые параллельно друг другу, что придает диффузной нервной системе более сложную форму, носящую название медуллярной.
Для сцифоидных медуз характерны скопления нервных клеток, близкие по строению с ганглиями. Если удалить все 8 краевых скоплений нервных клеток, то сокращения исчезнут, а раздражение будет вызывать только одноразовое сжатие колокола медузы. Нервная система кишечнополостных, организованная довольно просто с анатомической точки зрения, является основой довольно сложного и недостаточно исследованного поведения. У актиний вырабатывается устойчивое привыкание к однотипному механическому раздражению. Попытки обмануть актиний, подбрасывая пропитанную соком рыбы бумажку, приводят к выработке довольно устойчивого рефлекса. Через некоторое время актинии начинают отбрасывать ложную приманку, что говорит о существовании кратковременной памяти у кишечнополостных. Для кишечнополостных характерна и ассоциативная память. Если собрать актиний в природе и поместить в аквариум, то они расположатся в соответствии со своим исходным положением. Следовательно, каждая особь запоминает, в каком направлении было ориентировано ее ротовое отверстие.
К червеобразным относят животных, которые очень сильно различаются по размерам, форме тела, внутренней организации и образу жизни. Червя можно определить как билатерально-симметричное животное с мягким телом, длина которого в 2—3 раза и более превышает ширину. В отличие от двухслойной организации кишечнополостных у червей возникает третий зародышевый слой клеток — мезодерма. Она расположена между эктодермой (наружным слоем клеток) и энтодермой (внутренним слоем клеток). Этот признак характерен для всех других вышестоящих многоклеточных животных. Появление третьего слоя клеток определяет появление дифференцировки тела червей на внутренние органы. У большинства червей легко можно обнаружить выделительные органы и органы размножения.
Для червей характерна билатеральная симметрия. Это означает, что у червеобразных есть хорошо выраженный передний (головной) и хвостовой концы тела и сагиттальная плоскость, которая разделяет их на две более или менее симметричные половины тела. Наличие переднего конца тела, определяющего направление движения червя, привело к концентрации рецепторных нервных элементов в относительно небольшом участке тела — головной зоне. У большинства червей на головном конце тела имеется группа вкусовых клеток и светочувствительных элементов. Отростки этих клеток оканчиваются в головных скоплениях нейронов, носящих название ганглиев (рис. 7).
Конгломерат клеток, расположенных в головной части червя, может быть назван мозгом, поскольку выполняет характерные для него функции: получает, обрабатывает и передает к эффекторным органам информацию, поступающую от органов чувств. В качестве эффекторных органов у червей выступают мышечные клетки, которые позволяют осуществлять несложные движения ползания, плавания или демонстрируют простейшие ориентировочные рефлексы. Нужно отметить, что описанные выше особенности строения червей относятся к свободноживущим формам, тогда как у паразитических форм наблюдается редукция или полное отсутствие органов чувств, нервных клеток и органов движения.
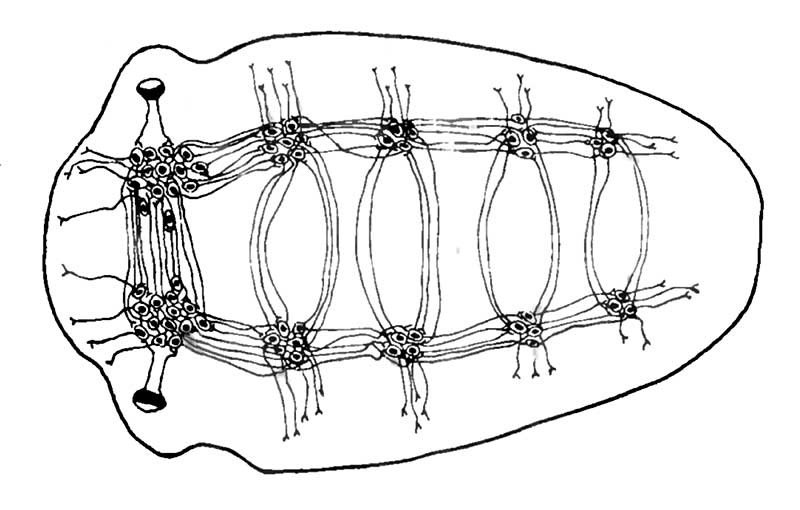
У плоского червя лестничная нервная система. В узлах лестницы находятся ганглии, которые соединены между собой поперечными комиссурами.
У плоских червей нервная система централизована менее всего. Два боковых нервных ствола тянутся у них вдоль всего тела от переднего конца к заднему (см. рис. 7). Боковые стволы соединены поперечными нервными пучками, а в зонах пересечений расположены тела нейронов. В общем виде нервная система плоских червей напоминает лестницу. Поэтому удачное название «лестничная нервная система» сохранилось до настоящего времени. Нервная система плоских червей расположена ближе к брюшной стороне тела, так как на ней сильнее развита мускулатура и она постоянно раздражается, соприкасаясь с субстратом, по которому ползет червь. К переднему концу тела размеры нервных стволов увеличиваются. В головной части червя расположен мозг, который состоит как из брюшных, так и из спинных скоплений нервных клеток.
Мозг плоских червей еще не играет принципиальной роли в поведении животного. При разрезании планарий на несколько кусков каждый из них двигается как целое животное, продолжает жить и регенерирует в целую особь. У некоторых морских червей головное скопление нервных клеток играет значительно более важную роль. При разрезании такого червя пополам его передняя половина тела двигается более правильно, чем задняя.
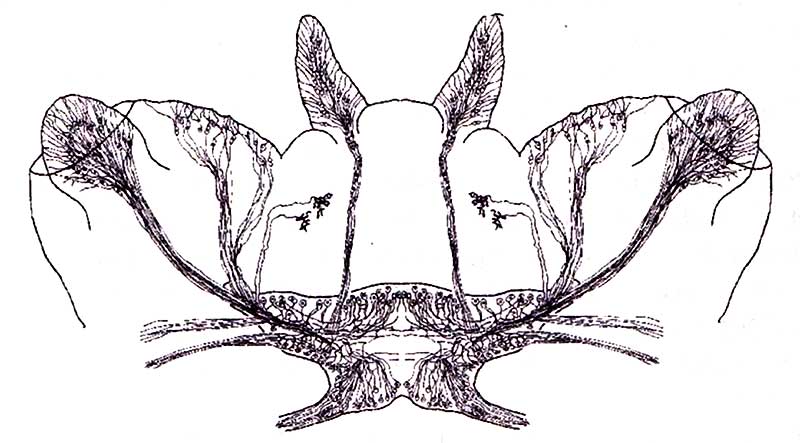
Значительно сложнее организована нервная система у кольчатых червей. Это обусловлено более сложным строением их тела и поведением свободноживущих форм. Тело кольчатых червей разделено на сегменты, называемые метамерами. Каждый из них содержит ряд органов, характерных для всего червя. Сегментация тела червя обусловлена развитием целома — внутреннего отсека, заполненного жидкостью и выстланного мезотелием — производным мезодермы. Кроме этого, кольчатые черви обладают сквозной пищеварительной системой, идущей от рта к анусу, и цепочкой нервных ганглиев, объединяющих все сегменты червя.
У кольчатых червей формируется ярко выраженная центральная нервная система. Клеточные тела нейронов сконцентрированы в отдельных сегментах так, что формируют два ряда нервных ганглиев. Ганглии каждого сегмента связаны поперечными волокнами между собой и продольными волокнами с ганглиями соседних сегментов. Ганглии сближены и образуют брюшную нервную цепочку. Головное скопление нервных клеток у кольчатых червей трансформировалось в настоящий мозг, расположенный над пищеводом (рис. 8). Ганглии имеют организацию, характерную для большинства беспозвоночных. Тела нервных клеток расположены на наружной поверхности, а отростки клеток и синаптические соединения — внутри ганглия. У кольчатых червей имеются многочисленные рецепторы, специализирующиеся на восприятии чувства равновесия, света и химических веществ. В примитивной форме присутствует соматосенсорная система, которая позволяет воспринимать сигналы, приходящие с поверхности тела червя. Кроме этого, кольчатые черви имеют специализированные рецепторы для осязания, ощущения давления и болевые рецепторы.
Многообразие рецепторов, большой мозг и весьма совершенная мускулатура позволяют червям вести разнообразную жизнь. Среди червей мы встречаем роющие, плавающие и ползающие формы. Они используют все виды растительной и животной пищи. Черви могут вести паразитический образ жизни, могут быть активными хищниками или вступать в симбиоз со свободноживущими животными. Некоторые полихеты (многощетинковые кольчецы) являются комменсалами морских звезд (используют хозяина как источник пищи). Полихеты находят своих хозяев в результате развитой хеморецепции. Они предпочитают здоровых и активных морских звезд. Если в Т-образном лабиринте им предложить чистую морскую воду и воду, в которой жила морская звезда, то они предпочтут последнюю. Если морская звезда была повреждена, то полихета предпочтет чистую морскую воду.
Для червей характерны все типы рефлексов, описанные для кишечнополостных. Однако организация нервной системы червей позволяет им проявлять более сложное поведение. Червеобразные способны к ассоциативному и условнорефлекторному обучению в Т-образном лабиринте. Обыкновенный дождевой червь использует 10—12 разнообразных приемов затаскивания опавшего листа к себе в норку. Он отказывается от попытки использовать в пищу лист, только перепробовав все возможные способы.
Это разнообразие поведения червей обусловлено усложнением организации нервной системы и появлением нервных клеток, специализирующихся на восприятии сигналов различной природы, которые позволяют червям вести свободноподвижный и хищнический образ жизни. Однако максимального развития нервная система беспозвоночных достигла у моллюсков и членистоногих.
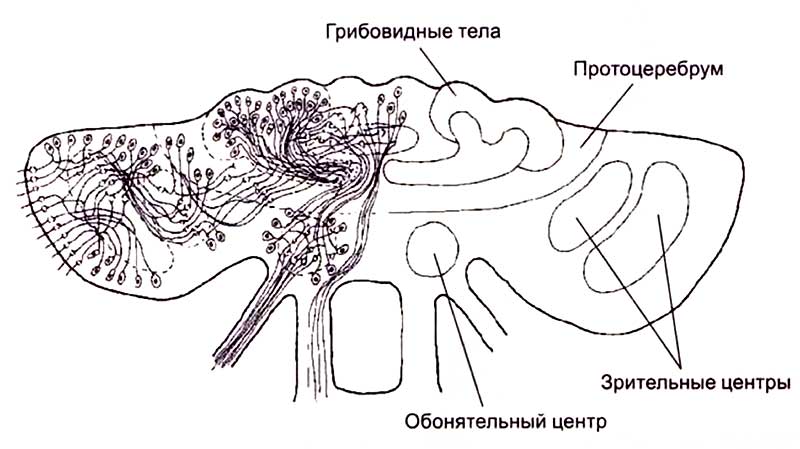
Для этой обширной группы животных характерно наличие по всей длине тела конечностей, которые иннервируются парными ганглиями, организованными в брюшную нервную цепочку. В головной части нервные клетки объединены в крупные ганглии, называемые мозгом. У этих животных головной мозг выполняет основные и незаменимые функции по интеграции жизнедеятельности организма. Развитие головного мозга членистоногих явилось результатом длительной цефализации нервной системы. Цефализация — увеличение размеров нервных центров в головной части животного — обусловлена концентрацией сенсорных органов на переднем конце тела червей, членистоногих и сходных с ними животных (рис. 9).
У членистоногих обнаружена сегментация мозга, вызванная появлением специализированных отделов, в которых происходит переработка информации, получаемой от сенсорных органов. У насекомых с хорошо развитым мозгом протоцеребрум получает информацию от глаз, дейтоцеребрум — от антенн с развитыми хеморецепторами, а тритоцеребрум — от передней части пищеварительного тракта. Наиболее важным результатом цефализации нервной системы является возникновение грибовидных тел, которые расположены в про-тоцеребруме членистоногих и некоторых высших червей. Грибовидные тела, по-видимому, участвуют в интеграции информации, поступающей от различных сенсорных систем. Они крайне сложно организованы. В шляпке грибовидного тела насекомых может располагаться от 3 до 12 слоев нейронов, размеры которых не превышают 2—3 мкм. Ножка грибовидного тела состоит из входящих и выходящих из шляпки волокон (рис. 10).
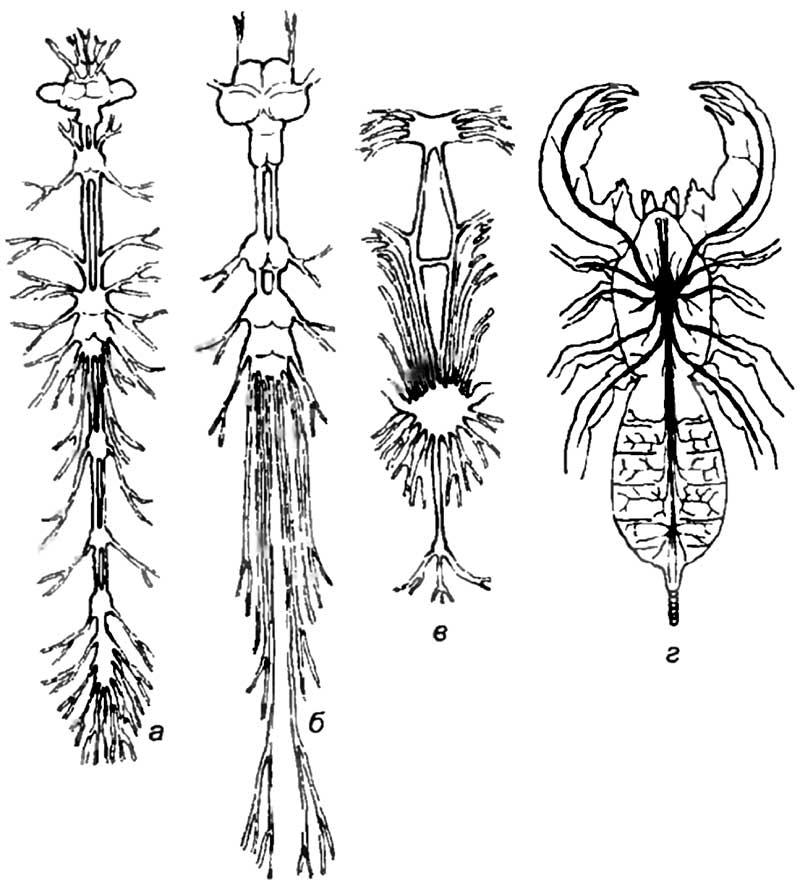
Грибовидные тела не выполняют функции непосредственной переработки информации, поступающей от слуховых, зрительных или химических рецепторов. В них происходит анализ уже первично обработанных сигналов от дистантных анализаторов и брюшной нервной цепочки. По сути дела грибовидные тела выполняют ассоциативные функции, позволяющие высшим беспозвоночным принимать решения на основе анализа информации, поступающей от всех систем организма.
Такая сложная организация мозга членистоногих используется для контроля самых разнообразных форм поведения. Примером может служить распознавание пищи падальной мухой. Она исследует пищу при помощи хеморецепторов, расположенных на дистальных сегментах ног. Контакт с потенциальной пищей вызывает прохождение нервных импульсов к подпищеводному ганглию и активацию его нейронов. В свою очередь эти нейроны возбуждают двигательные нейроны, которые стимулируют выворачивание хоботка и начало всасывающих движений. При разрушении мозговых ганглиев поиск пищи подавляется у аннелид и большинства насекомых. Необходимо учитывать, что рецепторы у насекомых намного чувствительнее, чем у других животных. Например, комары находят теплокровных животных, двигаясь по градиенту двуокиси углерода или аминокислот. Положительная реакция на аминокислоты выявляется у комаров при разведении 1:10 000 000.
На уровне головного мозга осуществляется главный контроль рефлексов и спонтанной активности ганглиев брюшной нервной цепочки. Он регулирует включение и выключение рефлекторных реакций низших центров, обладающих собственным аппаратом для организации конкретной деятельности.
Разрушение церебральных ганглиев у большинства беспозвоночных сопровождается гиперактивностью.
Не менее важным свойством сложного мозга является способность сохранять информацию. Большинство насекомых обладает хорошей памятью.
Наряду с мозгом и брюшной нервной цепочкой у членистоногих для управления организмом используются нейроэндокринные органы. Основные из них расположены позади головного мозга и соединены с ним посредством нервного ствола.
Нейрогормоны секретируются нервными окончаниями, локализованными в кардиальных телах нейронов мозга, или клетками самого кардиального тела. Вырабатываемые в этих органах гормоны необходимы для перестройки организма членистоногого во время метаморфоза — перехода от стадии личинки к взрослому животному (имаго).
а — личинка: б — комар после метаморфоза.
Во время метаморфоза происходит слияние грудных и брюшных ганглиев, что приводит к усилению цефализации нервной системы.
В этот момент значительным изменениям подвергается центральная нервная система насекомых (рис. 11). Так, например, у почти безногих личинок дровосека грудные узлы практически одинаковы по размерам с брюшными и незначительно отличаются по функциям. Среди этих ганглиев по размерам и функциям выделяется только головной мозг. У взрослого жука с развитыми ножными и крыловыми мышцами разница между узлами очень значительна. Во время метаморфоза происходит объединение нескольких грудных ганглиев личинки в один крупный ганглий взрослого жука.
Усложнение функций, связанных с полетом и движением конечностей, определяет увеличение размеров управляющих ганглиев. Необходимо отметить, что условия жизни личинки могут изменить поведение взрослого насекомого. Если личинки плодовой мушки дрозофилы были вскормлены на среде, содержавшей перечную мяту, то взрослые мухи обнаруживают положительный хемотаксис по отношению к этому растению. Эти данные свидетельствуют о сохранении нервной системой информации, полученной на личиночной стадии развития. Учитывая, что нервная система в период метаморфоза подвергается весьма значительным изменениям, запоминание «личиночного» опыта можно связать только с грибовидными телами или ассоциативными зонами церебральных ганглиев
Значение мозга для животных с таким сложным поведением, как у членистоногих, трудно переоценить. Даже в простых опытах по удалению головных ганглиев легко заметить глубокие изменения поведения. Жесткокрылые (жуки), раки и крабы после разрушения мозга начинают безостановочно ползать, однообразно очищать поверхность тела или совершать непрерывные жевательные движения. Стрекозы замирают на месте и не совершают никаких движений. Утрата головного мозга приводит к потере целесообразности поведения высших членистоногих, хотя автономные движения сохраняются. Сохранение способности осуществлять непрерывные движения при удаленном мозге определяется автономной активностью ганглиев брюшной нервной цепочки. Грибовидные тела мозга насекомых оказывают тормозящее влияние на активность движений. Локальное разрушение участков мозга вне грибовидных тел не влияет на активность движений, тогда как повреждение грибовидных тел вызывает усиление ходьбы и прыжков. Под контролем головных ганглиев находятся наиболее сложные поведенческие реакции, например пение насекомых. У сверчка контроль пения осуществляется грибовидными телами, центральным телом протоцеребрума и вторым грудным ганглием. Раздражая различные участки грибовидных тел и волокон, связывающих перечисленные ганглии, можно вызывать или прекращать песню сверчка. Кроме этого, стрекотание можно вызвать или затормозить, раздражая глаз, антенны, церки, тимпанальные органы или половой аппарат. Это показывает, что интеграция сигналов, приходящих от всех рецепторов, происходит в грибовидных телах, что и определяет тип песни сверчка.
Необходимо подчеркнуть, что органы чувств членистоногих хорошо развиты и могут быть специализированы так же, как и у позвоночных. Примером может служить способность совок (Heliothis armgera, Mythimna separata, Amphipyra perflua) к акустическому анализу ультразвуковых сигналов (Д.Н. Лапшин, 1995; Y. Хие et al., 1996). Эти бабочки способны к эхолокационной ориентации среди системы сферических преград. Они могут определять препятствие на расстоянии от 7 до 12,5 см и совершать разнообразные маневры их облета. Эти наблюдения показывают, что нервная система членистоногих достаточно развита для обработки и продолжительного хранения довольно сложной информации, которая необходима для формирования ассоциативного поведения.
Таким образом, у высших членистоногих мы сталкиваемся с новым качеством нервной системы. Мозг членистоногих выполняет настолько важные функции по интеграции поведения, что его утрата становится невосполнимой. Если удаление любого участка нервной системы плоских червей не приводит к гибели животного, го для членистоногих губительна утрата любого ганглия. Такая уязвимость нервной системы является результатом ее централизации и дифференциации строения головного мозга и ганглиев брюшной нервной цепочки. У насекомых мы встречаем крайне сложное поведение. Достаточно упомянуть широко распространенную заботу о потомстве и существование общественных насекомых. К ним относятся термиты, муравьи, осы, пчелы и ряд других насекомых, которые живут большими группами. Внутри таких сообществ различные особи выполняют специфические функции, что повлекло за собой внутривидовую специализацию нервной системы. Это вершина эволюции нервной системы беспозвоночных, поскольку выполнение общественных функций в других сообществах животных не приводит к морфологической трансформации нервной системы.
Среди беспозвоночных особое место занимают моллюски. Это довольно большая группа животных, включающая в себя несколько классов и около 80 000 видов. Рассмотрим основных представителей этой группы, коими являются брюхоногие (улитки), двустворчатые (мидии) и головоногие (кальмары и осьминоги) моллюски. У брюхоногих моллюсков тело обычно заключено в раковину, в большинстве своем они представляют собой малоподвижных животных; двустворчатые, как правило, вовсе неподвижные, прикрепленные организмы, а головоногие моллюски являются свободноживущими формами и весьма активными хищниками с практически полностью утраченной раковиной. Моллюски обладают крайне разнообразной нервной системой. У малоподвижных и примитивных форм она организована, как у плоских червей, а у головоногих моллюсков она наиболее сложная среди беспозвоночных (рис. 12—14).
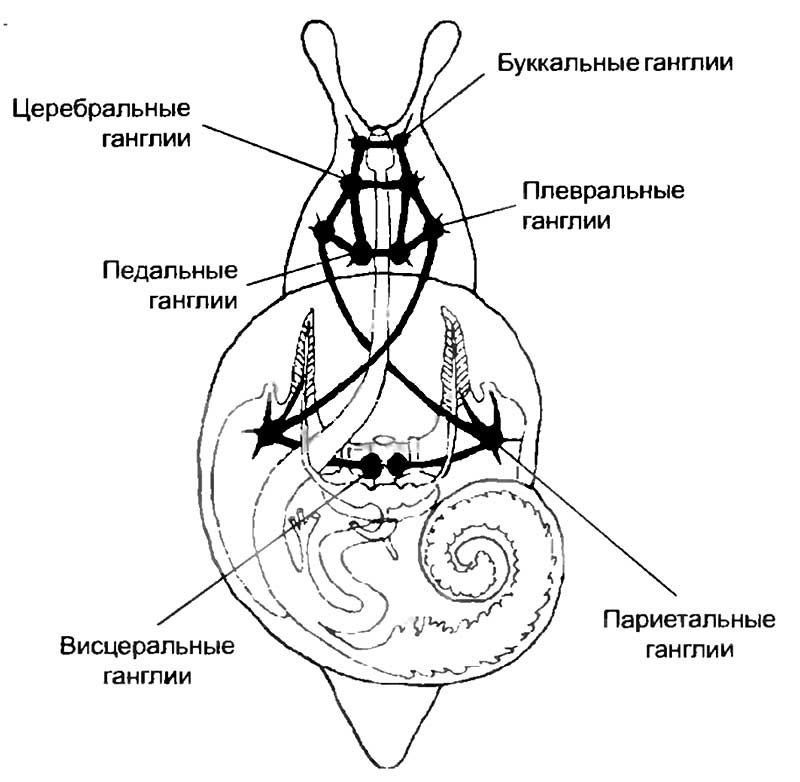
Наиболее типично организована нервная система у брюхоногих моллюсков (см. рис. 12). Головная часть нервной системы брюхоногих моллюсков состоит из четырех пар головных ганглиев, которые сгруппированы вокруг пищевода. Парные ганглии соединяются между собой комиссурами, а с ганглиями, расположенными вдоль оси тела, — коннективами, как у червей и членистоногих. Буккальный ганглий иннервирует рот и передний отдел пищеварительного канала, церебральный — глаза и щупальца, плевральный и педальный — «ногу». Отдельно расположены абдоминальный ганглий, иннервирующий органы висцерального мешка, и генитальный ганглий, лежащий вблизи органов размножения. Несмотря на простоту организации нервной системы брюхоногих моллюсков, они способны обучаться — вырабатывать условные рефлексы на внешние воздействия. Например, морской заяц (Aplysia) при длительной стимуляции жаберной пластинки и хвоста способен выработать кратковременный условный рефлекс, сохраняющийся несколько часов. Специальная программа обучения, приводящая к структурным изменениям нервной системы, позволяет возникнуть долговременной памяти, которая поддерживает выработанный навык в течение нескольких недель.
Нервная система головоногих моллюсков организована намного сложнее (см. рис. 13, 14). Это связано в первую очередь с потерей раковины и хищническим образом жизни. Большинство головоногих моллюсков — подвижные животные. Их движение осуществляется за счет выбрасывания воды через сифон по принципу реактивного движения. Это позволяет головоногим моллюскам активно охотиться и быстро избегать опасности. Головной мозг осьминога приближается к мозгу рыбы сходных размеров и состоит примерно из 150—180 млн клеток.
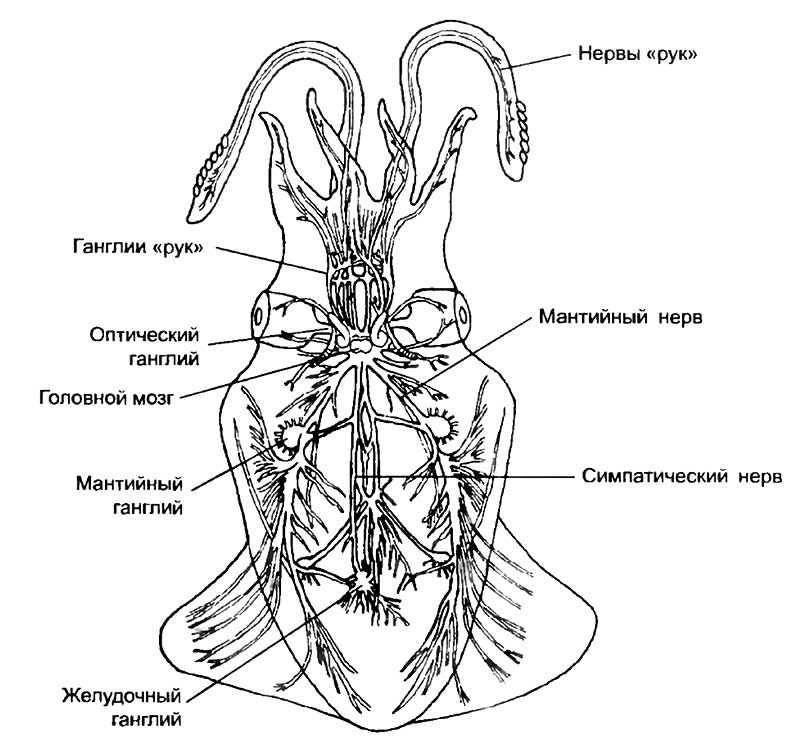
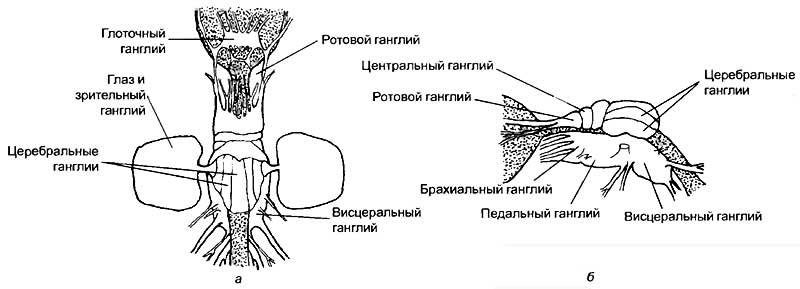
Головные ганглии нервной системы осьминога сосредоточены, как и у других моллюсков, вокруг пищевода (см. рис. 14). Ганглии увеличены, а поперечные комиссуры сильно укорочены. Слияние головных ганглиев приводит к формированию крупного мозга. Из органов чувств наибольшее развитие получили глаза. Нервные клетки специализированных зрительных долей мозга имеют различную морфологическую структуру, нехарактерную для остальных беспозвоночных, что указывает на использование зрительных центров в качестве ассоциативных областей. Крупными долями мозга осьминога являются брахиальные и педальные узлы, управляющие головными щупальцами и ротовой воронкой. Висцеральные ганглии иннервируют внутренности и содержат в себе автономный дыхательный центр с обособленной локализацией субцентров вдоха и выдоха. Буккальные ганглии определяют работу ротового аппарата. Сложность манипуляций, выполняемых щупальцами осьминога, не уступает способностям к манипулированию у хоботных (слонов) и человекообразных обезьян. Это достигается в результате интегративной работы центральной части мозга, контролирующей всё поведение осьминога. Однако центральные мозговые ганглии не являются однородным образованием.
Существует морфологическая дифференцировка, связанная с контролем конкретных функций. Первая пара центральных ганглиев контролирует движение «рук» осьминога во время поедания добычи. Вторая и третья пары центральных ганглиев через контроль педальных и брахиальных ганглиев управляют передвижением животного при помощи щупалец, осязанием и изменением направления при плавании. Во второй и третьей парах ганглиев находятся центры контроля окраски осьминога, которая постоянно изменяется, маскируя животное. При перерезке связей между этими ганглиями изменение окраски осьминога нарушается. Левая половина тела окрашивается иначе, чем правая. Для головоногих моллюсков контроль за окрашиванием тела имеет принципиально важное значение для реализации полового поведения. Перед спариванием у каракатиц самец становится полосатым, а самка, увидя его, принимает аналогичную пигментацию.
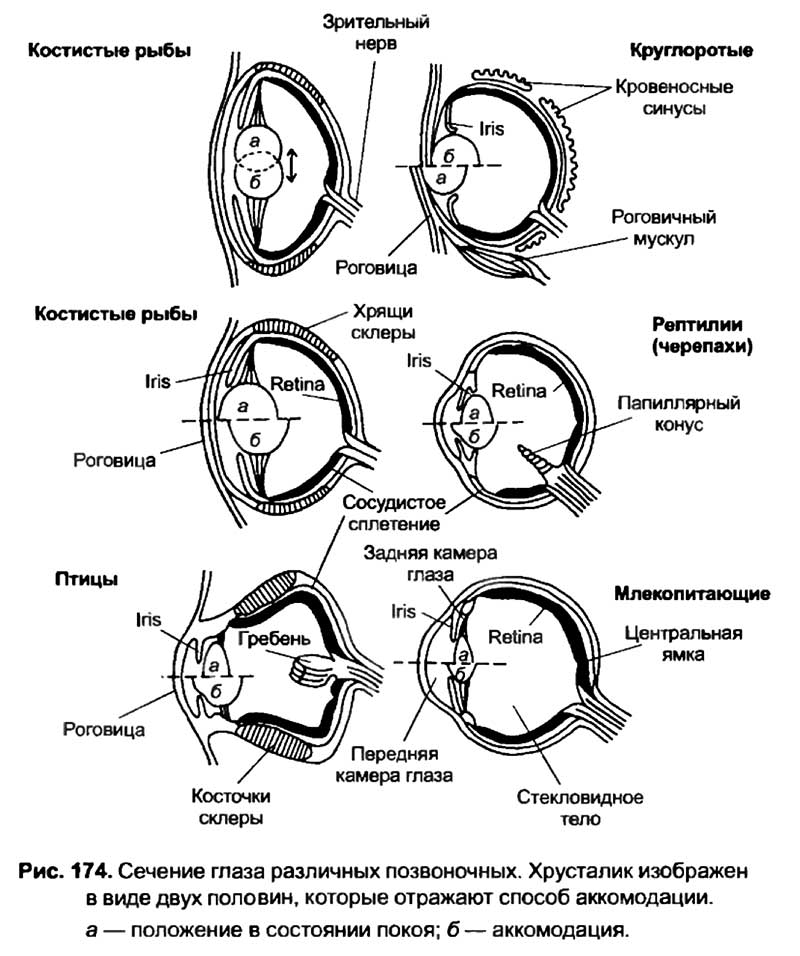
В жизни большинства головоногих моллюсков принципиальную роль играет зрение. Их глаза очень похожи на глаза позвоночных животных. У них есть роговица, радужная оболочка, сетчатка и хрусталик. У моллюсков наведение на резкость осуществляется за счет приближения или удаления хрусталика от сетчатки. Аналогичным образом происходит аккомодация хрусталика у анамний и змей (см. рис. 174). В области оптимальной зрительной чувствительности у каракатиц на один квадратный миллиметр приходится 150 000 зрительных рецепторов, тогда как у человека — только 65 000. Среди моллюсков наилучшее зрение имеет осьминог. Осьминог обладает некоторыми особенностями восприятия зрительных стимулов, поскольку он имеет горизонтально-щелевой зрачок и страдает астигматизмом. Эти анатомические особенности глаза приводят к тому, что осьминоги лучше различают вертикальные решетки, немного хуже — косые и хуже всего — горизонтальные (W.R.A. Muntz и J. Gwyther, 1989). Разрешающая способность глаз головоногих моллюсков сопоставима с таковой млекопитающих. Столь совершенное зрение позволяет осьминогам различать предметы различной формы при одинаковых размерах. У них после 3—4 предъявлений вырабатываются устойчивые рефлексы на сложные зрительные стимулы, подкрепляемые пищей. Головоногие способны к переделкам навыков, которые заключаются в использовании предварительно выработанных условных рефлексов для решения новых экспериментальных задач.
Наряду со зрением важным дистантным анализатором головоногих моллюсков является обоняние. Они могут различать очень низкие концентрации веществ, растворенных в воде. Осьминог способен обнаружить при помощи рецепторов на присосках хинин в концентрации, которая в 100 раз меньше ощущаемой человеком с помощью вкуса. Головоногие моллюски могут научиться различать большое количество зрительных образов, тактильных и химических раздражителей. Они умеют строить дома из камней, обладают хорошей зрительной памятью и легко обучаются.
Мозг головоногих моллюсков представляет собой наиболее сложную систему интеграции поведения среди беспозвоночных. Только самые высокоорганизованные насекомые обладают аналогичными способностями решать экспериментальные задачи, использовать память и предварительный индивидуальный опыт. Однако эти свойства нервной системы в наибольшей степени выражены у позвоночных животных.
Начиная изучать позвоночных животных, необходимо понять основные принципы их эволюции и историческую связь с беспозвоночными. Основным признаком позвоночных животных является наличие хорды — гибкой стержневидной структуры, которая проходит продольно через дорсальную [задне-верхнюю — спина, короче] часть тела. Она является эволюционным предшественником позвоночного столба и непосредственно связана с другими важными признаками — дорсальным полым нервным тяжом и глоточными жаберными щелями (рис. 15). В нервной системе позвоночных выделяют трубчатую центральную нервную систему и периферическую нервную систему, состоящую из ганглиев и нервов. Это разделение условно, так как периферическая нервная система в значительной степени состоит из отростков нейронов, расположенных в центральной нервной системе. Центральная нервная система подразделяется на спинной и головной мозг. Головной мозг расположен в мозговом черепе, а спинной мозг защищен невральными дугами позвонков, образующими вокруг него сегментированный чехол. В центральную нервную систему входят нервные волокна, идущие от различных чувствительных рецепторов. Волокна, выходящие из центральной нервной системы, передают сигналы от мозга к внутренним органам и мускулатуре животного.
а — прикрепленное кишечнополостное с радиальной симметрией;
б — свободноплавающее животное с радиальной симметрией;
в — придонное билатерально-симметричное животное с дорсальным жаберным аппаратом;
г — «ильное» животное, которое возникло из придонной формы в результате поворота тела на 90°;
д — примитивное хордовое с вторичной билатеральной симметрией тела;
е — свободноплавающее хордовое животное.
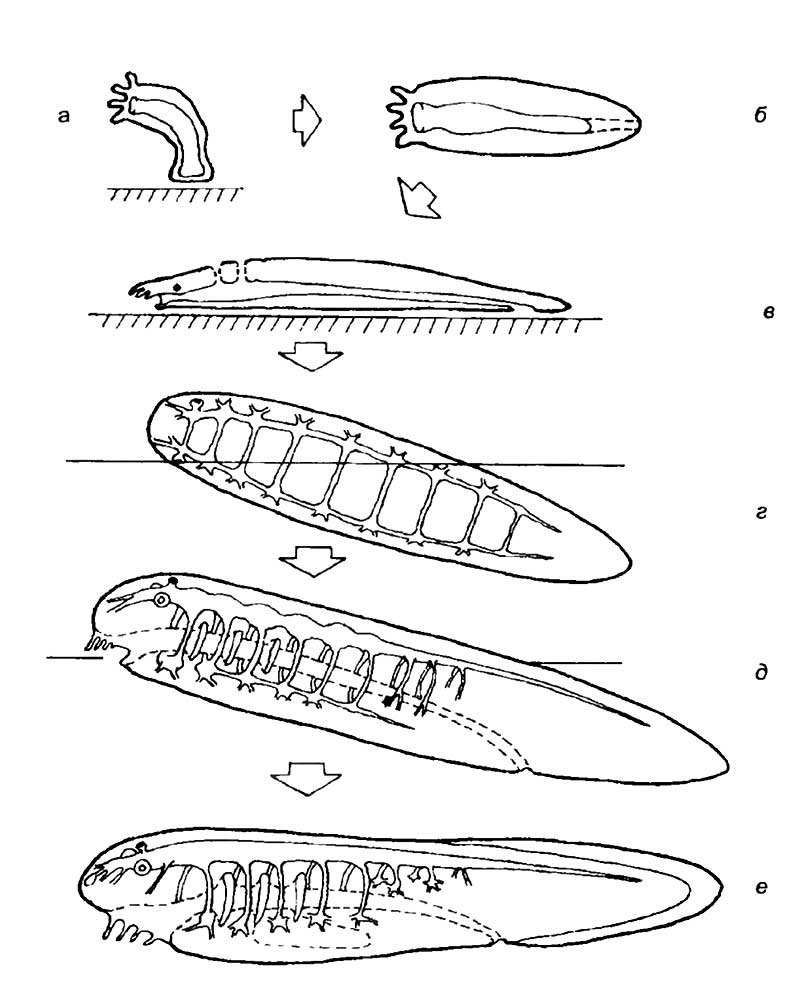
У наиболее низкоорганизованных хордовых можно заметить признаки организации нервной системы, сближающие их как с позвоночными, так и с беспозвоночными животными. Бытует несколько точек зрения на возникновение хордовых, которые имеют равные права на существование. Наиболее известны гипотезы возникновения хордовых от членистоногих, паукообразных, немертин, кольчатых червей, иглокожих, гребневиков и кишечнодышащих (В. Шимкевич, 1907; Р. Гессе, 1913; А.Н. Северцов, 1914; Н.А. Холодковский, 1914; В.Н. Беклемишев, 1944; Р.А Ливанов, 1955; Н. Kuhlenbeck, 1967; R. Jefferies и D. Levis, 1978; R. Norchcutt и C. Gans, 1983; А.В. Иванов, 1976, 1991; В.В. Малахов, 1977, 1980, 1991; Р. Кэрролл, 1992). Все эти теории с разной степенью доказательности отвечают на один вопрос о механизме возникновения трубчатой нервной системы позвоночных из ганглиозной нервной системы беспозвоночных. Рассмотрим наиболее логичную, с моей точки зрения, последовательность преобразования нервной системы в процессе эволюционного формирования хордовых. Допуская, что кишечнополостные являются безусловными предками основных групп животных, мы получим гипотетическое существо, изображенное на рис. 15,б, 16,а. Внутри животного проходит пищеварительный канал, который представляет собой гастро-васкулярную полость кишечнополостных, открытую спереди ротовым отверстием. У этого организма еще нет билатеральной симметрии, и он мог представлять собой какого-либо гидроида, перешедшего к свободноподвижному образу жизни. Он мог свободно перемещаться в трехмерной водной среде за счет сокращений щупалец, всего тела или пассивно, вместе с током воды. Основным результатом перехода к такому образу жизни стало появление окологлоточного нервного кольца.
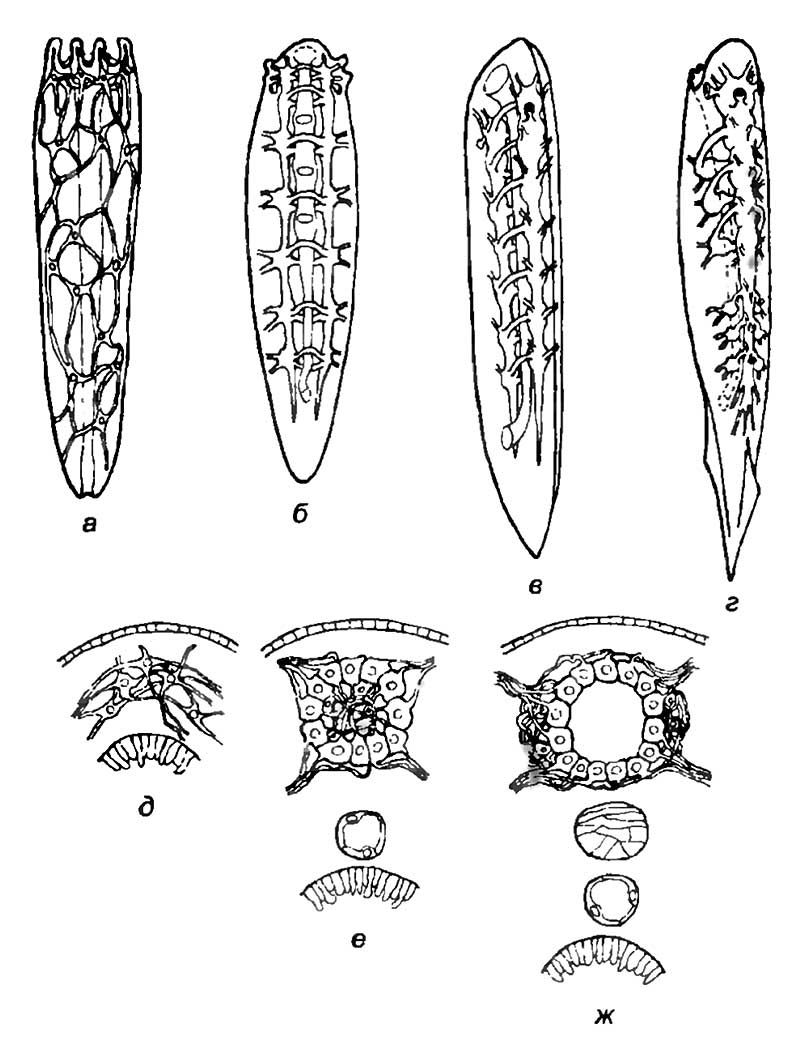
В верхнем ряду показаны четыре этапа централизации нервной системы. Нервная система вписана в контур тела гипотетических предков хордовых.
а — диффузная нервная система свободноплавающего животного с радиальной симметрией и окологлоточным нервным кольцом;
б — придонное червеподобное животное с плоским телом и двойной нервной цепочкой, расположенной по краям тела;
в — асимметричное «ильное» животное, возникшее из придонной формы после поворота тела на 90°; его нервная система состоит из двух нервных цепочек: дорсальной и вентральной. Дорсальная нервная цепочка увеличивается в размерах, а ганглии сливаются;
г — вторично-симметричное свободноплавающее животное с дорсальной нервной трубкой и сегментированной вентральной нервной цепочкой, из которой возникли вегетативные нервные ганглии. В нижнем ряду приведено схематическое изображение сечений тела гипотетических предковых форм хордовых:
д — радиально-симметричное животное;
е — придонное животное с билатеральной симметрией, мускулатурой, сегментированным телом, асимметричной кровеносной системой, кишечными отверстиями для дыхания и двумя тяжами нервных ганглиев;
ж — вторично-билатерально-симметричное животное с нервной трубкой и вентральными вегетативными ганглиями, возникшими в результате исчезновения продольных волокон «нижнего» тяжа ганглиев.
Животное с радиальной симметрией не нуждалось в усложнении организации, пока не перешло к придонному образу жизни. При передвижении по дну неизбежно должны были возникнуть спинная и брюшная поверхности тела и появиться признаки билатеральной симметрии. При переходе к активному движению по субстрату у предка хордовых возникла необходимость в усилении метаболизма. Это произошло за счет формирования на спинной стороне кишечной трубки щелей, которые выполняли роль жаберного аппарата (см. рис. 15,в). Замена радиальной симметрии на билатеральную с неизбежностью привела к возникновению парных нервных тяжей, идущих вдоль тела (см. рис. 15,г, 16,б). Вполне естественно, что ползание привело к формированию переднего (головного) скопления нервных клеток — головных ганглиев. Эти ганглии стали выполнять функции нервного центра, в котором концентрировались сенсорные и ассоциативные нейроны. У такого животного два нервных головных ганглия переходят в широко расставленные нервные тяжи, расположенные по бокам тела животного. В центре проходит пищеварительный канал, дорсальнее и вентральнее которого идут поперечные комиссуры, соединяющие левый и правый нервные тяжи (см. рис. 15,г, 16,б). Такое строение нервной системы характерно для многих плоских червей, хотя надо подчеркнуть, что описываемое животное является исключительно гипотетической моделью. При ползании по горизонтальной поверхности у животных самых различных систематических групп возникают сходные приспособления: пучки мышечных волокон, расположенные вдоль тела и имеющие сегментарное строение. Такая мускулатура необходима для создания волнообразных продольных движений тела, позволяющих двигаться головой вперед. Последовательно расположенные сегменты мускулатуры требуют согласования сокращений, что осуществляется посегментно расположенными латеральными ганглиями (см. рис. 16,б). Каждый мышечный сегмент приобрел собственный ганглий, соединенный с соседними ганглиями при помощи отростков нервных клеток. На этом этапе эволюции нервной системы произошло принципиальное событие, приведшее к дорсальному положению центральной нервной системы хордовых. Гипотетическое животное с двумя боковыми нервными тяжами развернулось на 90° — встало на «ребро». Одна боковая поверхность животного стала брюшной, а другая — спинной (см. рис. 15,г, 16,в). Вполне справедлив вопрос о причинах такого поворота. Реальной причиной может быть изменение структуры морского грунта, на котором обитало это гипотетическое животное.
Если поверхность дна постепенно покрывалась илом или животное обитало на границе интерстициали, то наиболее выгодно стало располагаться на ребре. В таком положении животное могло скрывать в илистых осадках большую часть тела, оставляя ротовое отверстие над поверхностью дна, и не затрачивало значительной энергии для маскировки и фиксации своего положения в грунте. Возможно и другое объяснение. Если эти животные обитали в прибрежных зонах моря, то становится актуальной проблема закрепления в грунте или быстрого погружения в него. Это могло быть связано с приливами и отливами, морскими течениями и подвижностью придонной воды мелководных морей.
В результате этого поворота один из боковых нервных тяжей стал брюшным, а другой — спинным. Животное оказалось асимметричным, так как новая ось симметрии совпала с горизонтальной плоскостью тела ползающего червеобразного предка. Поворот плоского червеобразного животного на ребро приводит к нескольким важным результатам. На верхнем (спинном) крае тела началось слияние ганглиев, составляющих один из тяжей нервной системы (см. рис. 15,д, 16,в). Это связано с тем, что при проталкивании тела в илистом грунте основное сопротивление приходится на переднюю часть тела животного, а задняя часть тела и хвост довольно пассивны. Поэтому координация движений должна быть максимальной в головной части животного, что и приводит к слиянию ганглиев в передней части дорсального нервного тяжа.
Увеличение именно дорсального нервного тяжа обусловлено тем, что он находился на границе ила и воды, в максимальном рецепторном контакте с внешней средой, тогда как вентральный тяж был по сути дела депривирован и изменил свои функции. На дорсальном тяже сохранились асимметричный глазок и тактильные рецепторы. Затем, после появления нервной трубки, дифференцировались боковые парные глаза и парные дистантные хеморецепторы. Вентральный тяж в головной зоне сохранил химическую и тактильную чувствительность, которая стала основой для развития органов чувств ротовой и жаберной области позвоночных.
Таким образом, центральная нервная система хордовых произошла из бокового (левого или правого) нервного ствола червеподобного животного, которое совершило поворот на ребро. Нервная система хордовых никогда не была в вентральном положении и является гомологом одного из боковых нервных стволов билатерально-симметричных беспозвоночных.
В результате объединения спинных ганглиев сформировалось протяженное скопление нервных клеток, не разделенное участками нервных волокон. Типичный ганглий беспозвоночного состоит из тел клеток, расположенных по его периферии, и центральной части, образованной из переплетения отростков нервных клеток — нейропиля (см. рис. 16, б, ё). При слиянии ганглиев произошло объединение нейропилей, тогда как сами нейроны остались на периферии нервного тяжа. Движение спинномозговой жидкости между отростками нейронов было слишком медленным для обеспечения метаболизма центральной нервной системы активного животного. Поэтому отростки вышли на наружную поверхность, освободив пространство — внутреннюю полость — для движения спинномозговой жидкости (см. рис. 16, ж). При этом симметричные комиссуры, идущие к брюшному нервному стволу, стали брюшными корешками спинного мозга, а дорсальные корешки возникли из чувствительных окончаний верхнего нервного тяжа. Следовательно, нервная трубка является гомологом слившихся ганглиев одного из нервных стволов беспозвоночных, внутри которых шизоцельно сформировалась полость — будущие мозговые желудочки и спинномозговой канал.
Совершенно другие изменения должны были произойти с брюшным нервным тяжом. Он не исчез и не редуцировался. У ползающего на ребре «ильного» животного брюшной нервный тяж сегментировался. Сегментация заключалась в том, что ганглии сохранили свое строение, но межганглионарные продольные связи стали иннервировать внутренние органы. Отростки нейронов, расположенных в вентральных ганглиях, стали преимущественно локализоваться в кишечном канале, стенках сосудов и выделительной системе, а не в соседних ганглиях. При этом ганглии брюшного нервного ствола не утрачивали комиссуральных связей со спинным нервным тяжом (см. рис. 15, д, е, 16, в, г). Вентральные головные скопления нервных клеток стали нейральной основой для совершенствования функций передней части пищеварительного канала и развития ротовой хеморецепции
Таким образом, ответ на поставленный вопрос о происхождении вегетативных ганглиев позвоночных довольно ясен. Вегетативные ганглии хордовых возникли в результате разделения одного из первично-латеральных стволов лестничной нервной системы беспозвоночных на отдельные ганглии, которые сохранили свои комиссуральные связи с противоположным нервным тяжом.
Следовательно, поворот на 90° уплощенного червеподобного животного привел к возникновению нервной системы, характерной для большинства хордовых: дорсальная нервная трубка и вентральные вегетативные ганглии. Ось симметрии изменилась, но это не повлекло за собой катастрофических последствий. Наоборот, исходя из этой модели, можно объяснить происхождение нервной трубки и вегетативных ганглиев. Нервная трубка хордовых гомологична одному из тяжей нервной системы уплощенных червеобразных беспозвоночных предков, тогда как вегетативные ганглии гомологичны второму боковому нервному тяжу этих животных.
Таким образом, рассмотрев становление нервной системы гипотетического предка хордовых, можно предположить основные события в филогенезе нервной системы, которые привели к формированию дорсальной нервной трубки и вегетативной нервной системы хордовых.
▢1. Самой древней предшественницей нервной системы хордовых была диффузная нервная сеть свободноплавающих кишечнополостных.
▢2. Появление парных нервных стволов произошло при переходе свободноплавающего кишечнополостного к придонному образу жизни. В результате возникли первая ось симметрии, брюшная и спинная стороны животного. Скорее всего таким животным было плоское червеобразное существо, ползавшее по дну.
▢3. Одиночный дорсальный нервный ствол возник после поворота червеобразного предка хордовых на 90°. Это могло произойти при переходе к ильному существованию на «ребре». В этом случае один из нервных тяжей оказался расположенным дорсально, а второй — вентрально. Из верхнего тяжа сформировалась центральная нервная система, а из нижнего — периферическая нервная система, иннервирующая внутренние органы.
▢4. Полость нервной трубки возникла после слияния ганглиев дорсального нервного ствола из нейропиля, который освободился от отростков нервных клеток. Они вышли на внешнюю поверхность слившихся ганглиев, сформировав слой белого вещества вокруг тел клеток. Причинами этих преобразований были сенсорно-привилегированное положение дорсального тяжа и необходимость усиления обменных процессов в мозге.
▢5. Сегментация центральной нервной системы хордовых возникла в соответствии с количеством мышечных сегментов, иннервируемых из двух нервных стволов червеобразного предка. Вентральные корешки спинного мозга сформировались из комиссур, а дорсальные (смешанные) нервы возникли из чувствительных и моторных нервов латеральной части тела червеобразного предка.
Описанное выше животное могло стать предком современных хордовых после повторного перехода к активному плаванию в трехмерной водной среде (см. рис. 15, е, 16, г). Для сохранения механической стабильности трубчатой нервной системы возникла хорда и изменился внешний вид животного. Следы описанных событий можно выявить при внимательном изучении личиночных и взрослых форм головохордовых и круглоротых.
Это большая группа морских животных, имеющих на стадии личинки выраженную спинную струну — хорду, над которой расположена нервная трубка (рис. 17, а). Личинки этих животных всегда свободноплавающие и всегда снабжены хвостом. Наиболее характерным представителем этой группы является асцидия. Свободноплавающие личинки асцидий по общему виду немного напоминают головастиков лягушек, которые обладают овальным телом и длинным, сжатым с боков хвостом. По оси хвоста идет хорда, представляющая собой с голбик студенистого вещества, одетый одним слоем клеток. Над хордой расположена нервная трубка, а по бокам — мышечные волокна. В теле дичинки нервная трубка утолщается, а в головной зоне приобретает вид пузырька. Это чувствительное образование является предшественником мозга и содержит в себе рецепторы. Личинка асцидии может воспринимать свет с помощью светочувствительного глазка и узнавать о направлении гравитационного поля благодаря наличию статоциста. Активно перемещающееся животное имеет глотку, сердце, околосердечную полость и выделительную систему, открывающуюся наружу одиночным отверстием. В таком состоянии большинство личинок находится только несколько часов. Затем они прикрепляются к какому-либо предмету при помощи специальных сосочков и превращаются во взрослое оседлое животное.
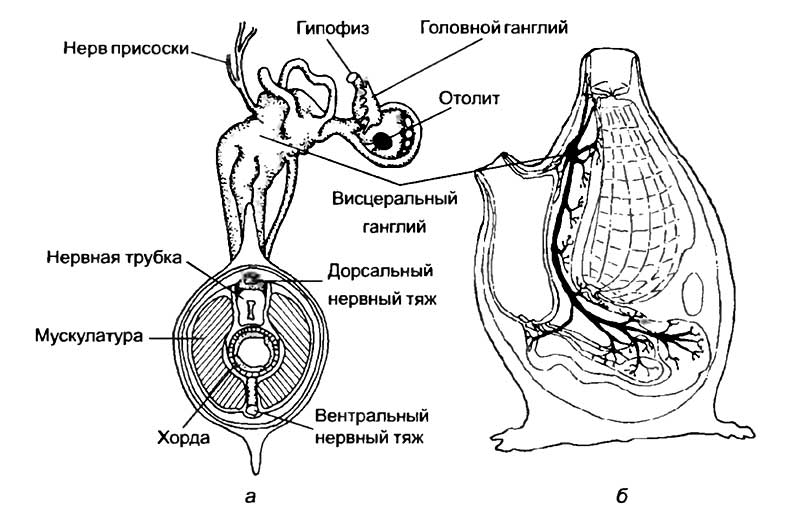
а — личинка асцидии, вид сверху и сечение; б — строение взрослой асцидии. У личинки нервная система представлена трубкой с асимметричным головным расширением, содержащим фоторецептор, и двумя нервными стволами. Один ствол лежит выше нервной трубки, а другой — ниже хорды. Для взрослой формы характерен один крупный нервный ганглий с отходящими от него сенсорными и моторными нервами.
Такое превращение носит название регрессивного метаморфоза. Он заключается в увеличении количества глоточных дыхательных отверстий, полном исчезновении хвоста и хорды. Наибольшим изменениям подвергается нервная система. Исчезают хвостовой отдел нервной трубки, глазок и статоцист. Вместо головного пузырька и туловищной нервной трубки формируется одиночный ганглий, характерный для нервной системы беспозвоночных (рис. 17, б), который лежит между ротовым и анальным отверстием. Он вытянут в спиннобрюшном направлении и формирует несколько нервных стволов. На брюшной стороне нервного узла расположена субнейральная железа, которую гомо-логизируют с гипофизом позвоночных. Таким образом, из активной свободноплавающей личинки с хорошо развитыми органами чувств, хордой и сложцо организованной нервной системой возникает сидячее взрослое животное, у которого органы, указывающие на родство с позвоночными, изменяются до неузнаваемости. Большую часть тела оседлой формы занимает пищеварительный тракт U-образной формы с обширной глоточной полостью Описанный жизненный цикл не является универсальным для всех оболочников. В некоторых примитивных родах этой группы хвостатой стадии развития нет, а у других развитие личинки происходит в родительском организме. Этому способствует гермафродитизм (двуполость) взрослых оболочников.
Личиночно-хордовые объединяют в себе признаки нервной системы позвоночных и беспозвоночных животных. Эта важная в эволюционном отношении группа показывает единство происхождения позвоночных и беспозвоночных животных, что имеет существенное значение для понимания исторического развития нервной системы. Следующей по уровню организации группой хордовых являются бесчерепные или головохордовые, которые обладают всеми основными признаками хордовых. Они имеют во взрослом состоянии хорошо развитые хорду, спинной мозг и жаберные щели (рис. 18). Единственным представителем группы является ланцетник — небольшое животное с прозрачным телом, которое заострено с обоих концов и сжато с боков. Нервная система ланцетника расположена над хордой, между мышцами правой и левой сторон тела.
Центральная нервная система имеет вид трубки, которая в поперечном разрезе представляет собой треугольник с полым центральным каналом. Сама трубка состоит преимущественно из продольных нервных волокон и тел клеток, сгруппированных вокруг центрального канала. Периферическая нервная система состоит из нервов, отходящих от центральной нервной системы. Они делятся на две группы: первая образована несимметричными парами нервов, отходящих от передней части нервной трубки, называемой мозгом, вторая состоит из большого числа спинных нервов, отходящих от спинного мозга (см. рис. 18,б). Первая пара головных нервов отходит от переднего края нервной трубки, а вторая — от его спинной стороны. Обе пары нервов имеют многочисленные нервные узлы, которые размещены в головной части ланцетника. Спинные нервы распределены посегментно, и соответственно расположению миомеров нервы правой и левой сторон отходят поочередно, а не друг против друга. В каждом сегменте с каждой стороны тела есть два нерва: спинной, отходящий одним корешком от верхней стороны спинного мозга, и брюшной, отходящий многочисленными отдельными волокнами. Спинные нервы идут к коже и к мышцам, являясь как чувствительными, так и двигательными. Брюшные нервы исключительно двигательные и идут к миомерам.
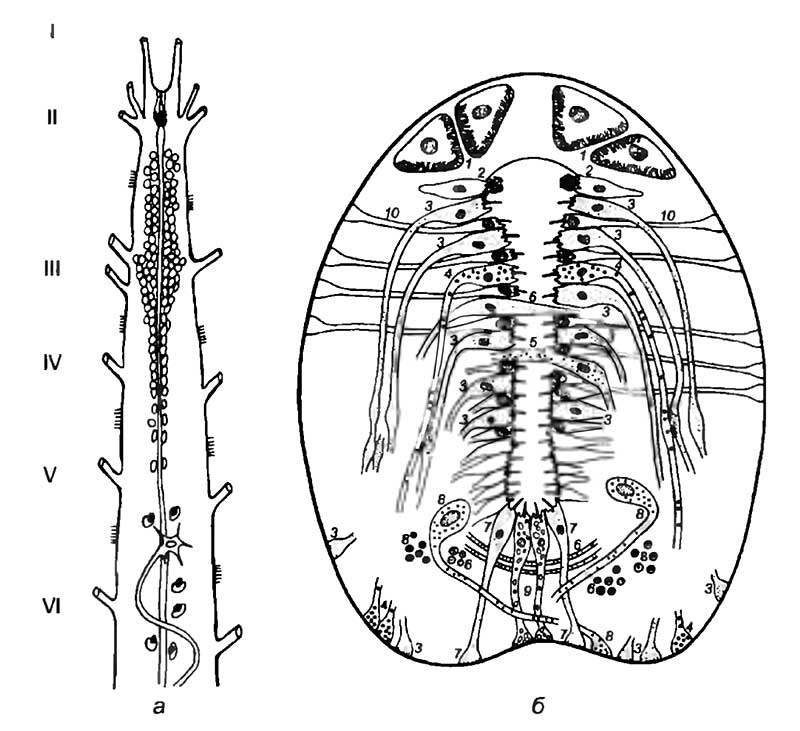
а — центральная нервная система, вид сверху: I-VI — головные нервы;
б — сечение тела взрослого ланцетника: 1 — клетки Жозефа (Joseph); 2 — ламеллярные фоторецепторные клетки; 3 — клетки, секретирующие катехоламины; 4 — нейроэндокринные клетки; 5 — пролактин-иммунореактивные клетки; 6 — кальцитонин-иммунореактивные клетки; 7 — катехоламинергические нейроэндокринные клетки; 8 — серотонинергические клетки; 9 — клетки воронки; 10 — клетки радиальной глии
В передней части нервной трубки центральный канал незначительно расширяется, образуя большую полость — желудочек передней части нервной трубки, а несколько позади него формируется спинное расширение. Это образование, как и вся нервная трубка, покрыто тонкой соединительнотканной оболочкой. Передний верхний отдел нервной трубки выдается небольшим полым мешком (асимметричная ямка Келликера), который соединяется с органом обоняния и носит название средней обонятельной доли. На его заднем конце с брюшной стороны есть небольшое углубление, называемое воронкой.
Органы чувств ланцетника представлены асимметричной обонятельной ямкой, открывающейся наружу на левой стороне головы. Этот хеморецепторный орган имеет полость, которая открывается через нейропор в центральный канал нервной трубки. Органы зрения ланцетника весьма своеобразны. В головной части они представлены непарным глазом, который выглядит как пигментное пятно. Небольшие светочувствительные клетки расположены в толще нервной трубки на всем ее протяжении. Каждая из них частично окружена пигментной клеткой. Такая пара клеток представляет собой отдельный простой глазок, способный улавливать различия в освещенности. Органы осязания представлены нейронами, иннервирующими ротовые щупальца, а орган вкуса — небольшой ямкой Гатчека, которая расположена на верхней стороне предротовой воронки.
К позвоночным относят обширную группу животных, включающую в себя круглоротых, рыб, амфибий, рептилий, птиц и млекопитающих. Обособление этой группы связано с тем, что все перечисленные животные обладают единым планом анатомического строения. У них имеются развитый позвоночник, череп, головной и спинной мозг. Именно внутри этой группы животных нервная система достигла своего максимального развития. Сложнее всего организован мозг млекопитающих и человека. Он обладает способностью не только получать, обрабатывать, хранить и передавать информацию, но и генерировать мысли о том, чего еще не было в природе.
Центральная нервная система позвоночных организована по общим принципам. В ней выделяют головной и спинной мозг, который расположен над позвоночником или хордой. Однако если головная часть ланцетника была намного меньше, чем спинная, то у большинства позвоночных это соотношение носит обратный характер. Головной мозг позвоночных в большинстве случаев намного больше спинного. Если принять величину спинного мозга за единицу, то относительная величина головного мозга будет следующая:
❏ пятнистая саламандра (хвостатые амфибии) — 0,9
❏ травяная лягушка (бесхвостые амфибии) — 1
❏ бык (млекопитающие) — 1,5
❏ карп (костные рыбы) — 2
❏ петух (птицы) — 2
❏ кролик (млекопитающие) — 2
❏ броненосец (млекопитающие) — 2,8
❏ ёж (млекопитающие) — 4
❏ кошка (млекопитающие) — 4
❏ летучая мышь (млекопитающие) — 6
❏ макака (млекопитающие) — 8
❏ дельфин (млекопитающие) — 18—23
❏ человек (млекопитающие) — 26
Интересно отметить, что у кошки и макаки приблизительно равная масса тела и спинного мозга (7,5 г). Масса же головного мозга сильно различается. У макаки он в среднем равен 60 г, а у кошки — 28—30 г. Таким же образом крупная собака, горилла и человек с близкими массами тела и размерами спинного мозга обладают различной средней массой головного мозга: 135, 430, 1350 г соответственно (рис. 19).
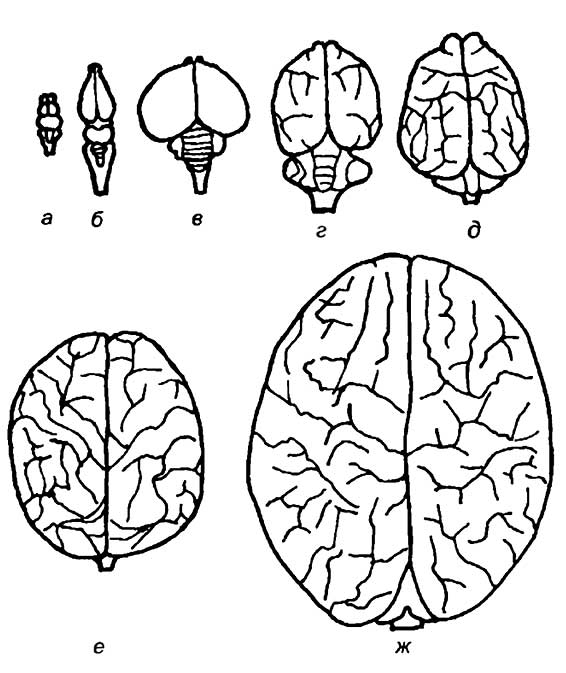
а — рыбы; б — рептилии; в — птицы; г — сумчатые млекопитающие; д — хищные; е — приматы; ж — человек.
Нетрудно понять причину этих различий. Масса спинного мозга и его размер непосредственно зависят от количества иннервируемых мышц и числа рецепторов, воспринимающих сигналы, идущие от тела животного. Чем больше размер тела животного, тем больше удельный вес спинного мозга в центральной нервной системе. По-видимому, зауроподы — самые крупные среди четвероногих травоядных динозавров — обладали спинным мозгом, во много раз большим по массе, чем головной мозг. С другой стороны, очевидно, что размер головного мозга не может определяться только периферическими анализаторами и иннервацией мускулатуры головы. Главная часть головного мозга позвоночных состоит из ассоциативных нейронов, которые не имеют никакой непосредственной связи с органами, воспринимающими раздражение, или с двигательными системами организма. Основная задача ассоциативных областей головного мозга заключается в интеграции всего разнообразия сигналов, приходящих из внешней среды и от внутренних систем организма. Анализ этой информации и использование предыдущего опыта определяют поведе-‘ние животного и его реакции на внешнее раздражение.
Таким образом, более совершенное развитие ассоциативных зон нервной системы является критерием приспособляемости и выживания конкретного организма. У высших позвоночных наиболее сложное поведение объясняется развитием ассоциативных центров головного мозга. Спинной мозг сам по себе вызывает лишь рефлекторные реакции, протекающие более или менее однообразно. Модификация этих реакций, их оттенки или полное подавление некоторых рефлексов — та нервная деятельность, которая сопровождается явлениями, называемыми памятью, рассудком и волей, — происходит в ассоциативных центрах головного мозга.
Для нервной системы позвоночных животных характерен ряд специфических характеристик. Центральная нервная система состоит из спинного и головного мозга. Головной мозг включает в себя пять основных отделов: передний, промежуточный, средний, задний и продолговатый мозг (рис. 20). Каждый из отделов головного мозга исторически связан с конкретными дистантными анализаторами — хеморецепторами, фоторецепторами, тактильными или слуховыми системами анализа внешней и внутренней среды организма. Как правило, конкретные рецепторы расположены на некотором расстоянии от мозга и соединены с ним посредством нервов. Нервы, относящиеся непосредственно к мозгу, носят название черепно-мозговых нервов. Всего их насчитывается 12 пар. Черепно-мозговые нервы разделяются на сенсорные, моторные и смешанные. Сенсорные нервы передают информацию от внешних и внутренних рецепторов в головной мозг, а моторные проводят эффекторные сигналы к мышцам.
Внутреннее строение центральной нервной системы характеризуется наличием полостей — мозговых желудочков, которые образуют систему сообщающихся цистерн, заполненных спинномозговой жидкостью (см. рис. 20). У низших позвоночных в переднем мозге расположена одна полость, которая носит название переднего мозгового желудочка (I). Для более высокоорганизованных позвоночных с парными полушариями переднего мозга характерно существование двух полостей (I, II), которые носят название латеральных желудочков (см. рис. 20, б). Строение переднего мозга многих млекопитающих и человека характеризуется наличием разветвленных латеральных желудочков переднего мозга. Желудочки формируют два или три рога, которые распространяются в лобную, височную и затылочную доли полушарий переднего мозга. Эти полости носят название рогов латеральных желудочков (см. рис. 20, в). Место схождения трех рогов носит название треугольника или звезды латеральных желудочков. Латеральные желудочки соединяются между собой и с III мозговым желудочком при помощи монроева отверстия.
а — организация, характерная для хрящевых и костных рыб,
б — для амфибий, рептилйй и птиц,
в — для млекопитающих.
I—IV — желудочки мозга. Передний мозг (telencepnalon), промежуточный мозг (diencephalon), средний мозг (mesencephalon), задний мозг (metencephalon), продолговатый мозг (myelencephalon).
III желудочек расположен на уровне промежуточного мозга и переходит в сильвиев водопровод. У низших позвоночных III желудочек и сильвиев водопровод образуют довольно широкую полость часто одинакового сечения на всем своем протяжении. Однако у рептилий и млекопитающих сильвиев водопровод значительно сужается. У человека его диаметр около 3—4 мм. Сильвиев водопровод переходит в IV желудочек, который лежит внутри заднего и продолговатого мозга. Стенки заднего и значительной части продолговатого мозга развернуты в латеральном направлении, что приводит к формированию широкой полости IV желудочка и истончению дорсальной стенки нервной трубки. IV желудочек сужается в каудальном направлении и переходит в центральный канал спинного мозга. Центральный канал проходит вдоль всего спинного мозга и иногда формирует расширение в поясничном отделе. Это расширение у млекопитающих носит название люмбального желудочка, а у птиц — люмбосакральной полости. Люмбосакральная полость часто заполняется у птиц желатинозной субстанцией, плохо проницаемой для спинномозговой жидкости. Надо отметить, что центральный канал спинного мозга у многих млекопитающих зарастает с возрастом на уровне плечевого пояса и грудного отдела позвоночника.
Внутри желудочков головного мозга и центрального канала спинного мозга проз гкает спинномозговая жидкость. Она образуется в сосудистых сплетениях за счет фильтрации плазмы крови. В полость желудочков из кровеносного русла проникают только вода и небольшое количество ионов. Сосудистых сплетений два (см. рис. 20). Одно расположено в латеральных желудочках и ростральной части III желудочка, а второе — в IV желудочке. Надо отметить, что у круглоротых существует сосудистое сплетение среднего мозга (plexus chorioideus mesencephali), которое выходит на наружную поверхность мозга. Переднее сосудистое сплетение обычно занимает 1/3 латеральных желудочков и 1/2 длины III желудочка. У млекопитающих оно проникает только в боковые рога латеральных желудочков. Заднее сосудистое сплетение обычно занимает большую часть крыши IV желудочка. У млекопитающих оно похоже на букву М. Интересно отметить, что у приматов ножки буквы М высовываются в специальные отверстия (отверстия Люшка) на наружную поверхность мозга около выхода слухового (VIII) нерва, а центральный изгиб сосудистого сплетения просовывается над мозговыми оболочками через асимметричное отверстие (апертура Мажанди). Эти отверстия являются своего рода страховочными клапанами, позволяющими быстро компенсировать резкие перепады давления спинномозговой жидкости. Сосудистое сплетение может иметь значительные морфологические различия у разных видов. Так, у круглоротых и двоякодышащих переднее сосудистое сплетение может покрывать своими складками практически всю дорсальную поверхность переднего, промежуточного и среднего мозга. Заднее сосудистое сплетение разрастается и закрывает собой как задний, так и продолговатый мозг. Мозг сверху выглядит как разросшаяся виноградная гроздь. У осетровых в развитом заднем сосудистом сплетении часто осуществляется кроветворение. Для многих костистых рыб характерен передний мозг, вывернутый внутренними поверхностями желудочков наружу. В этом случае сосудистое сплетение в переднем мозге практически отсутствует. Однако основная схема анатомического строения остается неизменной.
Как следует из названия группы, этих позвоночных объединяет отсутствие челюстного аппарата. Типичными представителями являются речная и ручьевая миноги. Кроме них в группу входят миксины — морские полупаразитические животные или падальщики. Круглоротые являются длиннотелыми первичноводными позвоночными, которые отличаются от рыб ротовой присоской без челюстей, непарным органом обоняния, отсутствием парных конечностей.
Нервная система миноги имеет ряд своеобразных черт, выделяющих ее из всех известных позвоночных животных. Спинной мозг миноги уплощен и напоминает по форме ленту. Его спинные корешки чередуются с брюшными и не соединяются с ними для образования смешанных нервных стволов. Симпатическая нервная система развита слабо и представлена несколькими небольшими узелками по бокам спинной аорты, в стенках сердца, в почках и половых железах. Первичный задний мозг миноги хорошо развит, а обширная полость IV желудочка занимает большую часть дорсальной поверхности этого отдела (рис. 21). По размерам он практически равен среднему, промежуточному и переднему вместе взятым. Интересно отметить небольшие размеры мозжечка, который выглядит, как небольшая поперечная лента у переднего края IV желудочка. Для животного, передвигающегося в трехмерной водной среде, такое строение мозжечка нехарактерно. Средний мозг, представленный парными зрительными долями, дифференцирован слабо. Его крыша открыта, как и полость IV желудочка заднего мозга. Поверхность крыши состоит из тонкого слоя недифференцированных нервных клеток и сосудистого сплетения, свешивающегося в полость сильвиева водопровода.
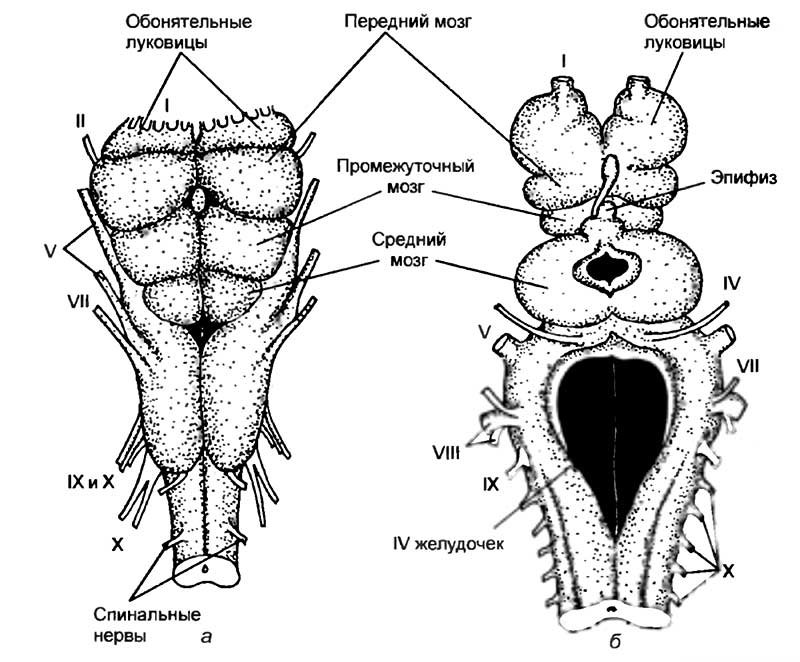
На верхней стороне промежуточного мозга находится два асимметричных утолщения — хабенулярные ганглии, из которых правое намного больше левого. В правый хабенулярный ганглий входит тонкий нерв от пинеального органа (эпифиза). Парапинеальный орган (теменной) посылает нерв в левый ганглий и по своему строению аналогичен пинеальному органу. Он содержит светочувствительные рецепторы, пигмент и несовершенный хрусталик. Это третий глаз миноги, позволяющий воспринимать изменения освещенности. Под эпифизом расположен вырост промежуточного мозга, связанный с левым хабенулярным ганглием. Он носит название парапинеального органа. Еще ниже расположен дорсальный мешок промежуточного мозга, который связан с обоими хабенулярными ганглиями. На нижней стороне промежуточного мозга расположен гипофиз. Передний мозг состоит из двух грушевидных выпячиваний. Парные задние утолщения являются собственно передним мозгом, а передние — обонятельными луковицами. У круглоротых орган обоняния непарный, хотя снабжается парным обонятельным нервом (I пара). У миног ноздря ведет через короткий носовой проход к обонятельному мешку, расположенному впереди головного мозга. От обонятельного мешка отходит кзади слепой вырост — назогипофизарный мешок, который прободает основание черепа и оканчивается под передним краем хорды. Стенка этого мешка прилежит к основанию промежуточного мозга и дает в развитии начало гипофизу. У мик-син этот мешок открывается в глотку. Осязательными органами служат нервные окончания в коже, а луковицеобразные группы нервных окончаний в ротовой полости являются органами вкуса. У миног двойная боковая линия, а лабиринт состоит из двух (у миноги) или одного (у миксины) полукружного канала. У миксин глаза рудиментарны, а миноги имеют крупный хрусталик и хорошо развитую толстую сетчатку. Важно отметить, что зрительные нервы (II пара) миног и миксин не образуют перекреста и направляются каждый к глазу со своей стороны. Для круглоротых характерно объединение IX и X пар нервов.
У рыб хорошо развиты органы движения, череп, челюстной и жаберный аппараты. Кроме жаберного аппарата, приспособленного для дыхания, у многих рыб есть плавательный пузырь, который в некоторых случаях может выполнять функции легкого или органа воздушного дыхания. У рыб парный орган обоняния только в виде исключения соединяется каналом с ротовой полостью. Питуитарный мешок, который хорошо развит у круглоротых, отсутствует, а гипофиз не имеет никакой связи с обонятельным мешком. В отличие от круглоротых лабиринт рыб имеет три полукружных канала. Среди хрящевых рыб наиболее интересной группой являются акулы. Различия в поведении акул коррелируют с морфологической организацией их мозга.
У активных хищников хорошо развито обоняние и высокоспециализирован передний мозг, который служит центром обработки обонятельных сигналов. У относительно неспециализированных форм размеры мозга сходны, но наиболее развиты ромбэнцефалон и мозжечок, которые являются координационным центром двигательной активности (рис. 22). Наибольшую вариабельность развития демонстрирует передний мозг акул и скатов. У пелагических акул, ведущих активный поиск пищи в верхних слоях воды, передний мозг обычно выглядит, как монополушарное образование (см. рис. 22,а). Для придонных форм акул характерно наличие парных полушарий (см. рис. 22,б). Особое строение имеет передний мозг у скатов. Парные обонятельные центры, которые формируются вокруг монополушарного переднего мозга, создают иллюзию его трехполушарной организации (см. рис. 22,в). Особняком стоит строение мозга химер, которые имеют чрезвычайно развитый продолговатый мозг (см. рис. 22,г).
У многих костистых рыб наиболее развиты зрительные отделы мозга. В качестве примера следует рассмотреть мозг костистой рыбы — карпа (рис. 23). При поверхностном вгляде на мозг карпа становится ясно, что основной структурой является средний мозг. В средний мозг приходит вторая пара черепномозговых нервов, идущих от глаз. Зрительные нервы у карпа перекрещиваются и идут на противоположную сторону: от правого глаза к левой половине среднего мозга, а от левого глаза — к правой. Мозжечок крупный по сравнению с другими отделами мозга и почти полностью закрывает полость IV желудочка. Передний и промежуточный мозг небольших размеров. Промежуточный мозг можно идентифицировать только по выходу эпифиза впереди парных выпячиваний среднего мозга (см. рис. 23). Нижняя часть промежуточного мозга формирует воронку и гипофиз. Из строения мозга совершенно ясно, что поведение карпа основано на хорошо развитом зрении и реализуется через координацию движений, которая осуществляется крупным мозжечком. С другой стороны, связанный с обонянием передний мозг развит плохо, что говорит о незначительном использовании хеморецепции. Для координации движений карпа большую роль играет анализ положения тела в пространстве, о чем свидетельствует развитый лабиринт с тремя полукружными каналами. Они позволяют карпу точно оценивать угловое ускорение движения. Органы линейного ускорения карпа представлены мешочками, содержащими крупные отолиты, которые являются кальцифицированными выделениями клеток.
Таким образом, строение мозга карпа отражает основные чувствительные механизмы, которые он использует для анализа окружающей среды. У рыб с другой экологией и способом добывания пищи строение мозга может существенно отличаться. Примером может служить мозг клюворылых (см. рис. 185).
У клюворылых электрическим органам, иннервируемым продолговатым мозгом, соответствуют особые «электрические полушария» мозжечка, которые по размерам превышают передний мозг. Они закрывают собой не только продолговатый мозг, но и все рострально лежащие отделы. У латимерии передний, промежуточный и средний мозг развит слабо. У нее относительно небольшой мозжечок и непропорционально крупные двигательные отделы продолговатого мозга (см. рис. 23, а).
а — длиннокрылая акула (Calcharhinus longimanus);
б — молодой (20 см) черноморский катран (Squalus acanthias);
в — гладкий скат (Raja batis);
г — европейская химера (Chimaera monstrosa).
I-X — черепно-мозговые нервы, зачернен IV желудочек.
Особенности строения тела некоторых рыб могут приподнести неожиданные нейроанатомические сюрпризы. Примером может служить строение центральной нервной системы рыбы-луны (см. рис. 23, 6). Этот вид из отряда скалозубообразных не имеет хвоста, питается зоопланктоном, откладывает до 300 млн икринок и ведет пассивный образ жизни. Отсутствие хвоста привело к укорочению спинного мозга, а неразвитость обоняния — к редукции переднего мозга. У рыбы-луны гипофиз лежит ростральнее переднего мозга, а самыми крупным отделом является средний мозг с развитыми зрительными нервами (см. рис. 23, б). Основной объем головного мозга занят нейрогормо-нальными и зрительными центрами, что полностью соответствует незатейлевой биологии рыбы-луны.
а — латимерия (Latimeria chalumnae);
б — рыба-луна (Mola mola);
в — карп (Cyprinus carpio);
г — бурый протоптер (Protopterus annectens).
I-X — черепно-мозговые нервы, зачернен IV желудочек, нл — нерв латеральной линии.
У двулегочников более сложная биология, которая включает в себя циклы спячки и активности, постройку гнезда, переход на различные типы дыхания и всеядность. У бурого протоптера мозг отражает многогранность его поведения (см. рис. 23, г). В его мозге хорошо развиты полушария переднего мозга, средний мозг и двигательные отделы продолговатого мозга. Поверхность IV желудочка протоптера практически закрыта сосудистым сплетением, которое формирует своеобразные «виноградные грозди». Аналогичное сплетение прикрывает промежуточный и передний мозг, но на рис. 23, г не показано.
Многие рыбы проявляют сложные формы поведения. Они узнают и защищают своего полового партнера, проявляют продолжительную заботу о потомстве, строят и охраняют свое гнездо, обладают способностью к обучению. Европейский лосось обучается выпрыгивать из воды и проскакивать через подвешенное в воздухе кольцо. Известны случаи обучения рыб нажиманию на рычаг для получения червяка. Реакция сохранялась месяцами, но без подкрепления быстро угасала.
Рыбы являются первичноводными животными, приспособленными к плаванию. Одна из наиболее важных черт их строения, накладывающая отпечаток на все их поведение, — отсутствие рычажных конечностей. Поэтому инструментальное поведение играет ограниченную роль в их жизни.
Амфибии внешне отличаются от рыб тем, что имеют рычажные конечности вместо парных плавников. В личиночном состоянии почти все амфибии дышат жабрами, которые у ряда видов сохраняются в течение всей жизни. У взрослых животных обычно имеются легкие. Личинки амфибий развиваются в водной среде и обладают хрящевым скелетом, тогда как большинство взрослых животных обитают на земле и имеют скелет из костной ткани. Переход от личиночной стадии к взрослому животному часто связан с выходом на сушу, что приводит к значительным изменениям скелета и внутренних органов. У лягушек исчезает хвост, развиваются легкие вместо жабр и формируются конечности. Этот процесс носит название метаморфоза. Некоторые земноводные становятся половозрелыми на личиночной стадии. Это явление лучше всего изучено у аксолотля и носит название неотении. Современные земноводные обладают гладкой кожей, которая легко пересыхает. Амфибии размножаются преимущественно в воде и не имеют тазовых почек. Поэтому они нуждаются во влажной среде и чаще всего обитают поблизости от водоемов. В эту группу входят бесхвостые (лягушки и жабы), хвостатые (саламандры и тритоны) и безногие амфибии — червяги (червеобразные роющие земноводные).
Нервная система амфибий имеет много сходных черт с рыбами, но обладает и рядом особенностей. Хвостатые и бесхвостые амфибии приобрели конечности, что повлекло за собой изменение организации спинного мозга. Спинной мозг лягушки короткий и оканчивается тонкой конечной ниточкой. В соответствии с числом позвонков у лягушки существует только 10 пар спинных нервов (рис. 24). II и III пары сливаются и образуют плечевое сплетение, проводящее нервы в передние конечности. VII, VIII, IX и X пары соединяются для образования крестцово-поясничного сплетения, которое иннервирует задние конечности. Такая специализация нервов спинного мозга впервые встречается у амфибий и отсутствует у рыб. Есть еще одно важное отличие в строении спинного мозга. У лягушки I спинной нерв выполняет функцию XII нерва и иннервирует мышцы языка.
Головной мозг лягушки отличается от мозга рыб очень небольшим мозжечком, более крупным передним мозгом и хорошо заметным промежуточным мозгом (см. рис. 24). Парные полушария переднего мозга имеют спереди крупные обонятельные луковицы, которые срастаются между собой по средней линии у бесхвостых амфибий. Промежуточный мозг расположен сразу за полушариями и имеет два выроста, соответствующих эпифизу и парафизу. Нижний мозговой придаток промежуточного мозга — гипофиз — хорошо развит. Средний мозг амфибий представляет собой наиболее крупную структуру нервной системы. У большинства амфибий он выглядит как парные полусферы, в которые входят зрительные тракты — продолжение зрительных нервов. Позади среднего мозга расположен крайне маленький мозжечок, который представляет собой небольшое утолщение передней стенки IV желудочка. Полость IV желудочка открыта. Все черепно-мозговые нервы хорошо заметны, но XII пара отходит от центральной нервной системы вне мозгового черепа и поэтому рассматривается как спинномозговой нерв. Добавочному (XI) нерву соответствует ветвь блуждающего нерва (X). Блуждающий нерв иннервирует внутренние органы и гортань (производное жаберного аппарата рыб). У амфибий парасимпатические ветви блуждающего нерва начинают иннервировать легкие. Языкоглоточный нерв (IX) иннервирует полость рта, глотку и язык. Он редко выходит из черепа отдельно, чаще всего сливается с блуждающим нервом.
У бесхвостых амфибий тройничный нерв (V) может сливаться после выхода из мозга с лицевым и слуховым нервами (VI—VIII). Зрительный нерв (II) образует на нижней стороне мозга срастание, напоминающее букву X, со сложным перекрестом волокон, тогда как у рыб это простое перекрещивание нервов. Обонятельный нерв (I) состоит из двух пучков волокон, которые у безногих амфибий разделяются и входят в мозг раздельно. Один из них идет от воздушного обонятельного мешка, а второй — от хеморецепторов якобсонова органа, который у амфибий выполняет функции органа водного обоняния.
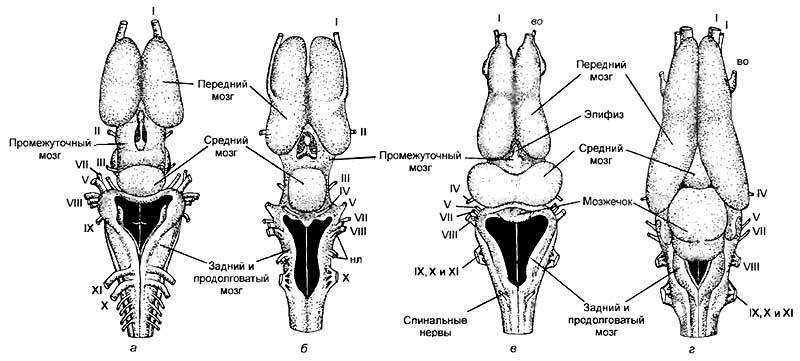
а — суринамская пипа (Рiра piра);
б — аксолотль (Ambystoma tigrinum);
в — травяная лягушка (Rana temporaria);
г — молодая (90 мм) червяга (Chtonerpeton viviparum).
I—XII — черепно-мозговые нервы, зачернен IV желудочек, во — вомероназальный нерв, нл — нерв латеральной линии.
Основные органы чувств амфибий изменены в соответствии с приспособлением к жизни в новой среде. У личинок и постоянно живущих в воде видов имеется сложная боковая линия, состоящая из нескольких рядов чувствительных луковиц, которая позволяет воспринимать колебания воды. У наземных видов эти органы заменены специализированными осязательными пятнами. Органами вкуса являются луковицеобразные тельца, разбросанные в слизистой оболочке нёба и языка. Носовые мешки, имеющие две связанные камеры, обладают способностью к хеморецепции как воздуха, так и воды благодаря расположенным в них рецепторным клеткам. Они открываются ноздрей наружу и специальным отверстием — хоаной — в ротовую полость. Так как вдыхание и выдыхание воздуха происходят обычно через ноздри, то носовая полость амфибий не только является обонятельным органом, но и входит в состав дыхательных путей.
Глаза амфибий позволяют различать цвета предметов и движущиеся объекты. Земноводные обладают хорошим зрением. Хрусталик их глаза сзади более выпуклый, чем спереди, и в обычном состоянии установлен для зрения вдаль. Аккомодация глаза достигается сокращением лежащей вокруг хрусталика мышцы, перемещающей его по направлению к роговице. У некоторых земноводных глаза невелики или рудиментарны и спрятаны под кожей.
Орган слуха включает среднее ухо, которое впервые встречаются у амфибий. Снаружи он ограничен барабанной перепонкой, за которой лежит большое пространство — барабанная полость, соединенная с ротовой полостью короткой евстахиевой трубой. В барабанной полости расположено стремя, передающее колебания барабанной перепонки на специальное окно внутреннего уха. Колебания воздуха вызывают движение барабанной перепонки, которое передается посредством стремени к эластичному окну внутреннего уха, обладающего механочувствительными клетками. Соединение евстахиевой трубы с ротовой полостью позволяет выравнивать давление воздуха в барабанной полости с атмосферным. Понятно, что такой орган слуха не отличается высокой чувствительностью и не позволяет воспринимать высокие звуки. Тем не менее возможность воспринимать колебания воздуха, а не воды впервые возникла у амфибий.
Симпатическая нервная система хвостатых амфибий состоит из большого симпатического нерва, простирающегося в виде цепи узелков, соединенных волокнами, от головы через туловище в хвостовой отдел тела. Там, где основные концы ребер раздвоены и охватывают позвоночную артерию, существует добавочный ствол симпатического нерва, связанный с большим симпатическим нервом.
Адаптация амфибий к наземному существованию повлекла за собой изменение поведения этих животных. Амфибии в большей степени, чем рыбы, отличаются индивидуальным поведением, которое координируется центральной нервной системой. Они проявляют различные формы заботы о потомстве и способность к обучению. Земноводные могут проходить простые лабиринты. В этом отношении жабы несколько превосходят лягушек, а тритоны и саламандры с такой задачей не справляются.
Переход к наземному образу жизни повлек за собой изменение организации центральной нервной системы и поведения земноводных. У наиболее высокоорганизованных амфибий становление поведения происходит в процессе индивидуального развития, Оно основано на врожденных стереотипах, которые тем не менее могут быть незначительно изменены в результате личного опыта конкретного животного. Это принципиальный шаг в эволюции нервной системы, поскольку способность к обучению и индивидуальному изменению поведения является мерой интеллектуальных способностей животных и человека.
При первом взгляде на пресмыкающихся становится заметно, что они отличаются от амфибий структурой кожи. Она всегда образует твердые роговые чешуи, иногда с кожными окостенениями внутри. Обычно конечностей две пары, хотя встречаются виды с одной передней парой или только с одной зачаточной парой конечностей. У змей и безногих ящериц ног нет вовсе. К ныне живущим рептилиям принадлежат ящерицы, хамелеоны, змеи, черепахи и крокодилы. Эта обширная группа животных освоила огромные территории благодаря своей независимости от воды в период размножения.
Нервная система у рептилий развита значительно лучше, чем у амфибий. Спинной мозг имеет цилиндрическую форму с плечевым и поясничным утолщениями, соответствующими конечностями. Сзади спинной мозг продолжается, постепенно сужаясь, в начало хвостового отдела позвоночника
Головной мозг у рептилий, особенно у крокодилов и черепах, более широк и массивен, чем у амфибий. Для мозга рептилий характерно изгибание вниз в области продолговатого мозга и стремление крупных полушарий переднего мозга надвинуться на лежащие позади промежуточный и средний мозг (рис. 25). Полушария переднего мозга без резких границ переходят в обонятельные луковицы, от которых начинаются обонятельные нервы (I). Увеличение размеров полушарий переднего мозга связано с появлением зачатков коры — одного или нескольких слоев клеток, расположенных под поверхностью мозга. В этой зоне переднего мозга осуществляется вторичный анализ информации, поступающей от органов обоняния. Промежуточный мозг почти совсем закрыт сверху передним мозгом, так что на поверхность высовывается только париетальный (парапинеальный) орган. У многих ящериц этот орган сохраняет строение глаза. Как и другие позвоночные, рептилии имеют воронку, переходящую в гипофиз. Средний мозг представлен в виде хорошо развитого двухолмия, которое выполняет функции зрительных центров. Однако у змей зрительное двухолмие несколько редуцируется и впервые появляются небольшие парные задние бугорки — среднемозговые центры слуха (у рыб они обслуживали органы боковой линии). Двигательные отделы мозга рептилий представляют собой крупное образование с почти закрытым IV желудочком. Он прикрыт сверху мозжечком, который может иметь вид полоски, треугольника, овала или круга, как у крокодилов. У последних можно выделить центральный и два боковых отдела мозжечка.
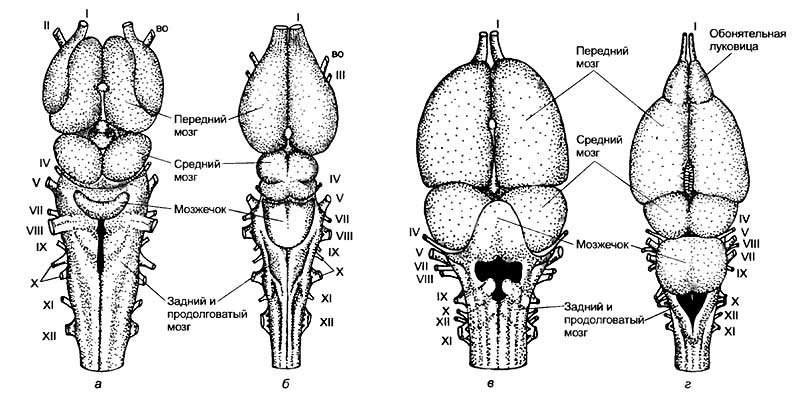
а — геккон (Gekko декко);
б — сетчатый питон (Python reticularis);
в — хамелеон (Chamaeleo lateralis);
г — красноухая черепаха (Pseudemys skripta).
I—XII — черепно-мозговые нервы, зачернен IV желудочек, во — вомероназальный нерв.
Черепно-мозговые нервы представлены 11 парами, которые преимущественно обособлены друг от друга. Нервы, часто сливающиеся с тройничным и блуждающим нервами, выходят из мозга рептилий по отдельности. Только у змей лицевой нерв иногда тесно связан с узлом тройничного нерва. Обонятельный нерв (I) снабжает своими ветвями обонятельный мешок и якобсонов орган. Зрительные нервы (II) образуют в основании черепа перекрест, называемый хиазмой. При этом у яшериц волокна одного зрительного нерва проходят сквозь щель другого. Лицевой нерв (VII) представляет небольшую слаборазвитую ветвь.
Блуждающий нерв (X) является одним из наиболее длинных черепно-мозговых нервов. Это обусловлено тем, что сердце, желудок и легкие сильно удалены от головы. К блуждающему нерву прилежит добавочный нерв (XI), а подъязычный нерв (XII), начинаясь несколькими корешками от передней части спинного мозга, выходит из черепа через одно или несколько специальных отверстий.
Симпатическая нервная система представлена у рептилий симпатическим нервом, который в голове образует соединение с ветвями многих черепных нервов. У крокодилов в шейном отделе он разделяется на два тяжа. Один из них лежит в позвоночном канале, образованном раздвоенными основаниями ребер, а другой — на нижней поверхности шейных позвонков. У змей шейная и туловищная части симпатического нерва слабо развиты.
Органы чувств рептилий обладают рядом уникальных особенностей, что связано с ороговением покровов. Несмотря на ороговение, в чешуйках кожи многих рептилий найдены осязательные пятна, а под чешуйками существует развитая сеть осязательных телец с нервными окончаниями. Огромное значение в качестве осязательного органа имеет язык, который у многих змей и ящериц постоянно находится в движении. Вкусовыми органами служат луковицеобразные тельца в слизистой стенке языка и нёба. Орган обоняния у рептилий представлен двумя системами: основной (обонятельный эпителий носовых мешков) и добавочной (якобсонов орган). Носовая полость парная, сообщается с ротовой и глоточной полостями через хоаны. У ящериц и змей часто встречается парная полость, лежащая книзу от носовой и открывающаяся особым отверстием в ротовую полость. На дне полости расположены хемочувствительные карманы — якобсонов орган, в который рептилии вкладывают раздвоенный на конце язык. Слуховой аппарат рептилий состоит из внутреннего и среднего, а иногда и из зачатков наружного уха. Лабиринт более развит, чем у амфибий, а у крокодилов уже появляются признаки улитки. Глаза рептилий обычно несколько крупнее по отношению к голове, чем у амфибий. Глазное яблоко шаровидной формы, а вокруг роговицы у многих ящериц и черепах формируется склеротикальное кольцо из костных пластинок. Аккомодация у большинства рептилий достигается при помощи мышцы, расположенной вокруг хрусталика. Сокращение этого мускула сжимает хрусталик, делая его толще, и тем самым дает возможность рассматривать близко расположенные предметы. Только у змей аккомодация происходит за счет перемещения хрусталика, как у амфибий. Кроме того, у некоторых змей имеются терморецепторы очень высокой чувствительности. Они могут улавливать инфракрасные волны от мыши, температура тела которой выше, чем температура окружающей среды, с расстояния. 15 см. Так как терморецепторы расположены в лицевых ямках, гремучие змеи могут определять направление источника излучения.
В колбочках рептилий и птиц находятся специальные цветные масляные капли, которые сужают кривые спектральной чувствительности рецепторов и уменьшают их взаимное перекрытие. Масляные капли в 2—5 раз увеличивают число различимых цветовых оттенков у этих животных. Многие черепахи обладают хорошим цветовым зрением. В сетчатке одного из исследованных видов (Pseudemys scripta) было найдено несколько типов фоторецепторов, отвечающих за восприятие цвета и освещенности: палочки, красные колбочки, красно-зеленые двойные колбочки, одиночные красные и зеленые колбочки. Все эти фоторецепторы работают в различных условиях освещенности фона и объекта, что позволяет черепахам использовать 4 механизма цветовосприятия.
Поведение рептилий построено на врожденных реакциях, которые значительно индивидуализируются в процессе жизни. Надо отметить, что продолжительность приобретения индивидуального опыта у рептилий довольно велика. Известно, что крупные черепахи живут 150—250 лет, змеи и ящерицы — 7—25 лет, крокодилы — 70—80 лет. У рептилий очень большую роль играет обучение. Многие из них легко приручаются и быстро осваивают лабиринт. Черепахи могут узнать фигуру, нарисованную сплошной линией, даже если она очерчена пунктиром или повернута на небольшой угол. Для ящериц характерно некоторое прогнозирование развития событий. Это позволяет им преследовать добычу уже после того, как она скрывается из виду. Интересно отметить, что водяные ужи, обученные правильно проходить Т-образный лабиринт, после линьки все забывают. Аналогично ведет себя шипохвост, который, привыкнув к новой территории, забывает ее после кратковременного охлаждения.
Для рептилий характерна иерархическая организация компактных групп. Доминирующей игуане уступают место не только на своей территории, но также и на соседних, когда она направляется на охоту в общие нейтральные зоны. Иерархия у гигантских черепах Галапагосских островов выражается в строго установленном порядке следования друг за другом во время миграции. Групповая организация рептилий является одной из сложных форм общественного поведения, которая была достигнута у холоднокровных животных.
Птицы отличаются от остальных позвоночных тем, что они имеют крылья, покрыты перьями и способны поддерживать постоянную температуру тела. Это яйцекладущие животные, которые по строению центральной нервной системы весьма сходны с рептилиями. Птицы являются высокоспециализированными животными, приспособленными к обитанию в воздушной среде, и почти каждая часть их организма адаптирована к столь необычным условиям. Приспособлениями к полету являются хорошая теплоизоляция, оперение, превращение передних конечностей в крылья, видоизменение грудины и плечевого пояса для крепления больших мышц крыла, а тазового пояса с задними конечностями — для поддержания массы тела на земле, а также усовершенствование дыхательной системы, которая обеспечивает наивысшую температуру тела среди всех животных. Кроме этого, у птиц отсутствуют зубы, утрачены левая дуга аорты и правый яичник с яйцеводом. Этих особенностей строения достаточно для получения ясного представления о том, что птицы являются высокоспециализированной группой позвоночных. С другой стороны, организация современных птиц в наиболее существенных чертах довольно однообразна.
Строение мозга у птиц значительно менее разнообразно, чем у рыб, амфибий и рептилий. Головной мозг заполняет полость черепа и имеет округлую форму при крайне малой длине (рис. 26). Самый крупный из отделов головного мозга передний мозг. Он состоит из двух полушарий, которые имеют гладкую поверхность или слабо обозначенные продольные углубления (см. рис. 26). Эти углубления не являются бороздами, а отражают границы слоев неостриа-тума. Только у попугаев можно заметить небольшое морфологическое обособление височной доли полушарий. Основной объем полушарий занимают подкорковые ядра, тогда как кора имеет рудиментарное строение и занимает небольшую часть верхней стенки мозга. Роль коры млекопитающих выполняет неостриатум, который развивается из гомологичного участка стенки полушария. Полушария переднего мозга простираются назад до контакта с мозжечком. Поэтому парные выпячивания среднего мозга до такой степени сдвинуты наружу, что лежат по бокам.
Следствием разрастания переднего мозга назад, а мозжечка вперед является то, что промежуточного мозга снаружи совсем не видно, хотя его можно определить по выросту эпифиза. Эпифиз у птиц развит незначительно, тогда как гипофиз имеет достаточно крупные размеры. Средний мозг сильно развит, но из-за своего бокового расположения имеет нехарактерную внешнюю морфологию. Мозжечок состоит из массивной средней части (червя), пересекаемой обычно 9 извилинами, и двух небольших боковых долей, которые гомологичны клочку мозжечка млекопитающих.
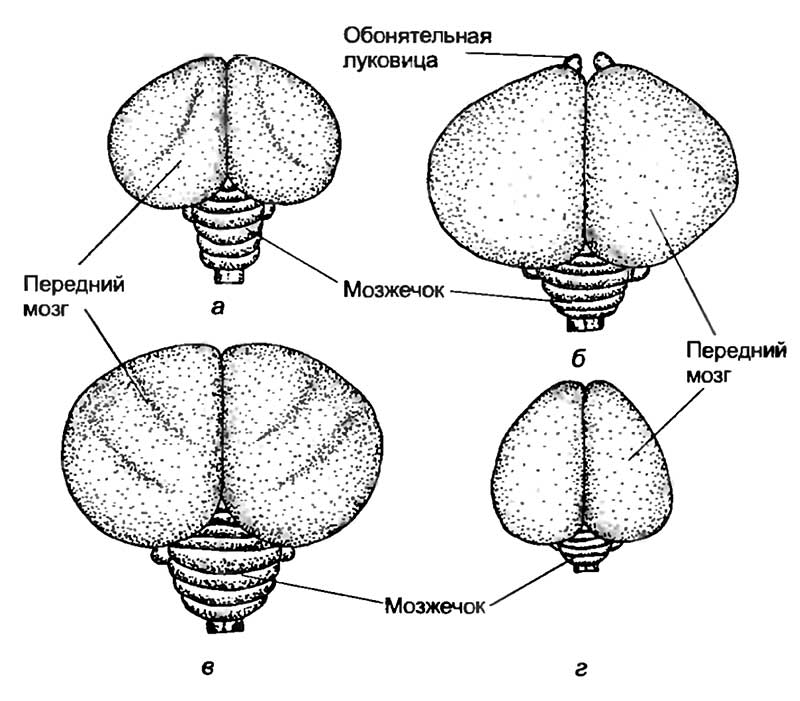
а — обыкновенная индейка (Meleagris gallopavo);
б — серый гусь (Anser anser);
в — бразильский ябиру (Jabiru myeterica);
г — обыкновенная зеленушка (Chloris chloris).
Продолговатый мозг относительно мал, его нижняя сторона в еще большей степени, чем у рептилий, образует изгиб вниз, а в области промежуточного мозга наблюдается изгибание вверх. Спинной мсзг птиц очень хорошо развит. Он образует большие утолщения в плечевой и поясничной области, откуда отходят нервы передних и задних конечностей. В поясничном утолщении верхняя стенка спинного мозга расходится и центральный канал расширяется в ромбовидный синус, покрытый сверху только мозговыми оболочками. Спинномозговые нервы соединяются у птиц корешками еще в канале позвоночного столба и выходят между дугами или через отверстия слившихся позвонков. В плечевом сплетении обычно участвуют 4 нерва, реже — 3. а у страусов — только 1 нерв. Для управления задними конечностями у птиц существует два сплетения: поясничное и седалищное, или крестцовое. Поясничное сплетение обычно состоит из 3, а у страусов может включать и 5 нервов. Седалищное сплетение у всех птиц состоит из 4 крестцовых нервов.
Черепно-мозговые нервы птиц представлены 12 парами, которые начинаются отдельно. Обонятельный нерв (1) идет от нижней поверхности обонятельной доли, которая спереди и снизу примыкает к полушариям переднего мозга. Зрительные нервы (II) после хиазмы плавно переходят в средний мозг, а блуждающий нерв, как и у рептилий, идет из черепа далеко назад, иннервируя сердце, легкие, пищевод и желудок. В отличие от рептилий добавочный нерв (XI) представлен тонкой веточкой блуждающего нерва, а подъязычный (XII) нерв имеет корешки, отходящие как от продолговатого, так и от спинного мозга.
Симпатическая нервная система птиц построена по общему для всех позвоночных принципу. Однако отличительной чертой ее строения у птиц является расположение шейной части большого симпатического нерва, часто называемого пограничным стволом. Он лежит в канале, образованном двукорневыми началами поперечных отростков. Участок этого нерва прилежит к сонным артериям, а далее кзади симпатический нерв находится в канале, образуемом головками и бугорками ребер, и только в поясничной области он лежит свободно.
Строение органов чувств птицы несколько отличается от такового остальных животных. Осязание многих птиц (кулики, утки) сосредоточено в челюстном аппарате, в котором расположены специализированные механорецепторы (тельца Гранди и Хербста). Осязательные тельца расположены в коже у основания крупных перьев и сконцентрированы в восковице, которая покрывает основание клюва. Совы обладают особыми осязательными перьями, расположенными вокруг клюва. У дятлов, дроздов, попугаев и глухарей осязательные функции выполняет язык, весьма богатый нервными окончаниями. У птиц с хорошо развитым языком вкусовые рецепторы играют роль в выборе пищи. Зрение птиц очень хорошо развито. Глаз способен к аккомодации и обладает рядом морфологических особенностей, позволяющих птицам эффективно ориентироваться, успешно охотиться и издалека определять качество пищи. У птиц отлично развит слух. Орган слуха состоит из внутреннего, среднего и зачатков наружного уха. Птицы могут воспринимать сигналы различной частоты. В этом отношении они намного превосходят многих млекопитающих.
Поведение птиц крайне сложно и разнообразно. Они заботятся о потомстве, совершают перелеты на расстояния в тысячи километров, имеют сложное брачное поведение, строят гнезда или специальные устройства для выведения птенцов, обучают молодых птиц и сохраняют способности к обучению на протяжении всей жизни. Птицы обладают хорошей памятью и могут осуществлять несложную инструментальную деятельность. Они легко обучаются решать простые задачи и подражать человеческому голосу. «Слова», произносимые птицами, являются эмоциональным символом настроения, в котором они их запомнили, а не результатом рассудочной деятельности.
В отличие от всех рассмотренных выше групп животных птицы любят играть друг с другом или с различными предметами. Мозг птиц обеспечивает возможность предвидения вероятных событий. Они рассчитывают свое поведение на несколько ходов вперед, что уже характерно для высших млекопитающих и человека.
Млекопитающие являются высшими из позвоночных животных. Они теплокровны и выкармливают своих детенышей молоком. Все млекопитающие всю жизнь дышат воздухом при помощи легких, хотя многие из них — водные животные. Тело млекопитающих в большинстве случаев четко разделяется на голову, туловище, хвост и четыре конечности. У китообразных и сиреновых развиты только передние конечности, а у некоторых приматов и человека отсутствует хвост. В группу млекопитающих входят однопроходные (утконос и ехидна), сумчатые (американские опоссумы, бандикуты, вомбаты и кенгуру) и плацентарные звери (грызуны, китообразные, копытные, хищные, насекомоядные, приматы и т.д.). Масса головного мозга у млекопитающих очень различается (табл. 1).
| Вид | Масса головного мозга, г |
|---|---|
| Мышь (грызуны) | 0,4 |
| Крыса (грызуны) | 1,6 |
| Морская свинка (грызуны) | 4,8 |
| Кролик (зайцеобразные) | 9,3 |
| Кошка (хищные) | 31,0 |
| Собака (хищные) | 65,0 |
| Макака-резус (приматы) | 88,0 |
| Корова (парнокопытные) | 340,0 |
| Шимпанзе (приматы) | 355,0 |
| Горилла(приматы) | 940,0 |
| Человек (Homo sapiens) | 1350,0 |
| Дельфин (китообразные) | 1550,0 |
| Слон (хоботные) | 6075,0 |
| Финвал (китообразные) | 7200,0 |
| Кашалот (китообразные) | 9200,0 |
Как видно из табл. 1, мозг млекопитающих может различаться по массе в 10 000 раз. Это разнообразие массы мозга соответствует разнообразию размеров животных. Однако каждый вид весьма неоднороден. Он состоит из подвидов и пород, которые имеют существенные различия как по размерам тела, гак и по объему мозга.
Примером таких различий могут служить собаки. Их мозг может различаться по массе в несколько раз. Самая маленькая масса мозга собаки, которая была описана в литературе, составляла 41,5 г, а самая большая — 212 г (Н.И. Петропавловский, 1904). Ниже представлена внутривидовая изменчивость массы мозга собак (в граммах):
❏ Минимальная масса мозга — 41,5
❏ Максимальная масса мозга — 212,0
❏ Средняя масса мозга беспородных собак — 73,2
❏ Средняя масса мозга породистых собак — 92,8
❏ Средняя масса мозга сук — 76,9
❏ Средняя масса мозга кобелей — 85,0
Весьма показательно отношение массы мозга и массы тела. Минимальное отношение 0,2 выявлено у крупных собак, масса тела которых составляет 28—44 кг. Для собак массой менее 2,5 кг оно увеличивается до 2,9. Следовательно, масса мозга по отношению к массе тела уменьшается с увеличением размеров животного. Эта закономерность не наблюдается в период развития щенков. У щенков и взрослых собак одной и той же породы отношение массы мозга к массе тела практически одинаково. Это отличает индивидуальное развитие мозга собак от такового обезьян и человека. У обезьян и человека плод рождается с большим мозгом и маленькой массой тела, а затем масса тела увеличивается намного быстрее, чем масса мозга. Головной мозг подвержен половому диморфизму. У самок масса мозга обычно на 10—12 г меньше, чем у самцов. Тем не менее суки обычно мельче, чем кобели, что заметно влияет на относительную массу мозга и тела. У сук относительная масса мозга больше, чем у кобелей.
Весьма любопытны различия породистых и беспородных собак. Средняя масса мозга в выборках беспородных собак колеблется от 42 до 118 г, а в группе породистых — от 41,5 до 212 г. Искусственно выведенные породы имеют большую амплитуду изменчивости головного мозга, чем беспородные собаки. По-видимому, у одичавшей в Австралии дикой собаки Динго мозг имеет еще большую однородность по массе, чем у беспородных городских собак. Возврат к природным формам уменьшает искусственно выведенное разнообразие. Надо отметить, что у породистых собак относительная масса мозга почти в два раза меньше, чем у беспородных.
Центральная нервная система у млекопитающих развита больше, чем у какой-либо другой группы животных. Основными отличиями строения мозга млекопитающих от мозга рептилий и птиц являются крупные парные полушария переднего мозга, которые у высших зверей формируют борозды и извилины.
Крупные полушария млекопитающих исторически возникли из боковой части стенки переднего мозга рептилий в результате разрастания поверхностного скопления клеток, которое носит название плаща или коры. У высших млекопитающих новая кора состоит из 6 четко выделяемых слоев, толщина которых у высших приматов доходит до 100 клеток. Общее число нейронов в коре переднего мозга человека составляет 11 млрд клеток. В коре млекопитающих продублированы почти все основные функции нервной системы на уровне заднего, среднего, промежуточного и переднего мозга.
Кора подразделяется на поля — зоны, в которых представлен конкретный периферический анализатор или моторная система. Примером может служить затылочная кора человека, которая ответственна за зрение и дублирует функции среднего мозга низших позвоночных. Наличие коры позволяет млекопитающим точнее и детальнее анализировать сигналы, приходящие из внешней среды и от внутренних органов, что лежит в основе сложного поведения и мышления человека. Однако только улучшение анализа рецепторных сигналов не может быть достаточным условием для становления сложного поведения. Необходима глубокая интеграция всей информации, поступающей в мозг. Эти функции выполняет кора за счет связей между расположенными в ней нейронами. Подсчитано, что на каждый мотонейрон приходится до 2000 вставочных нейронов, которые позволяют согласовывать активность отдельных клеток. Таким образом, увеличение размеров коры и формирование ее слоистой структуры позволили повысить качество обработки поступающей в мозг информации и интегрировать активность отдельных его структур. На этом морфологическом субстрате возникло новое качество мозга — способность к синтетической деятельности.
Полушария переднего мозга млекопитающих разрастаются вбок и кзади, покрывая собой промежуточный мозг, часто также средний мозг, а у человека и продолговатый мозг (рис. 27). Полушария переднего мозга имеют неправильную овальную форму и более или менее расширены сзади. У многих млекопитающих, принадлежащих к самым разнообразным группам (насекомоядные, грызуны, зайцеобразные), полушария гладкие, без признаков борозд и извилин. У большинства высших млекопитающих в коре формируются борозды и извилины (см. рис. 27). Однако извилины бывают в различной степени развиты даже в пределах одного отряда. Низшие приматы практически лишены борозд и извилин, тогда как у высших приматов их строение значительно сложнее (рис. 28). С увеличением размеров полушарий непосредственно связано появление мозолистого тела — обширной зоны волокон, соединяющих левое и правое полушария. Это образование характерно для мозга млекопитающих. Промежуточный мозг полностью скрыт полушариями переднего мозга. В нем хорошо развит эпифиз, а воронка, как и у всех позвоночных, завершается крупным гипофизом. Он выполняет преимущественно нейроэндокринные функции, в частности для регуляции сезонной, суточной и половой активности млекопитающих.
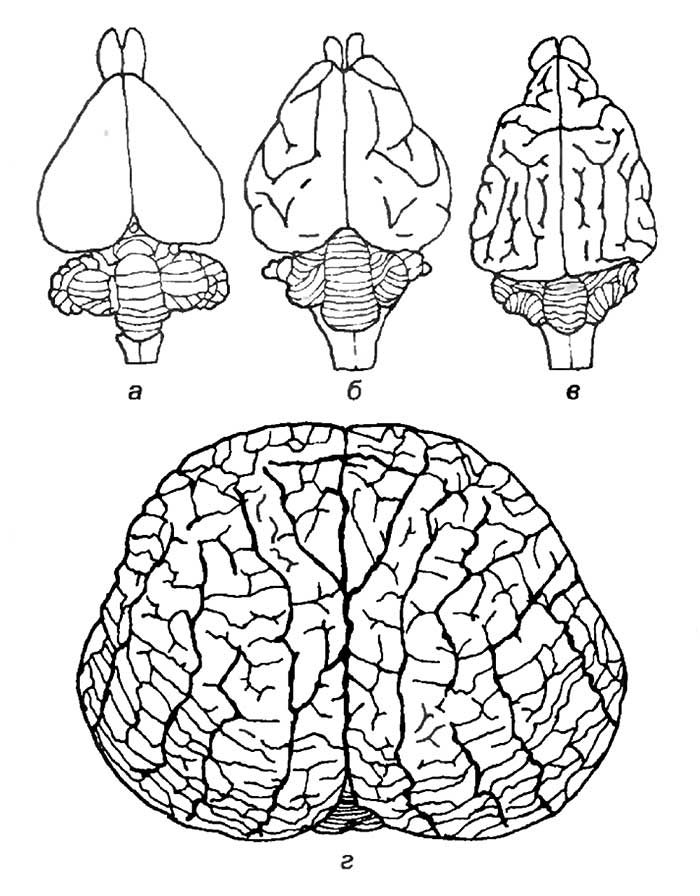
а — кролик; б — кенгуру; в — волк; г — дельфин (афалина).
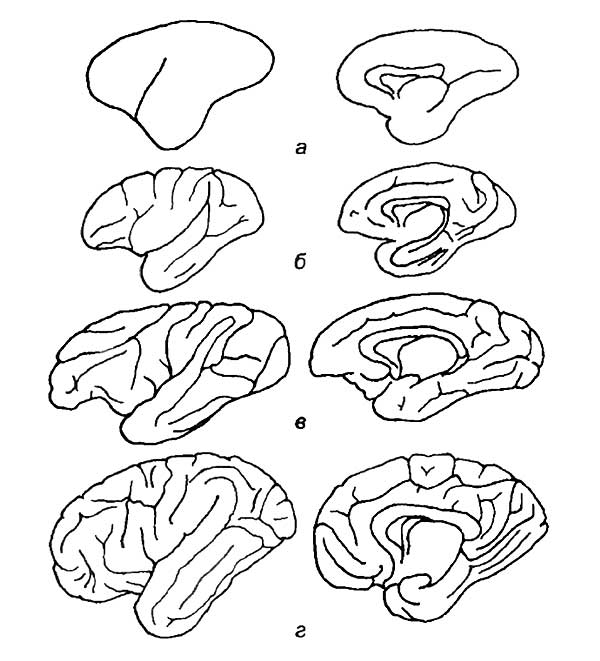
а — лемур (Lemur variegatus);
б — капуцин (Cebus apella);
в — павиан {Cynopithecus niger);
г — орангутанг (Рондо pygmaeus).
Средний мозг представляет собой плотное небольшое тело в виде двух пар возвышений, которые носят название четверохолмия. У рептилий и амфибий крыша среднего мозга состоит из двух возвышений, связанных с сетчаткой. У млекопитающих они отчасти сохраняют свои функции и расположены в передней зоне четверохолмия. Появление задних бугорков среднего мозга связано с развитием слухового аппарата млекопитающих. Задние бугорки среднего мозга являются слуховыми центрами и отражают уровень развития слуховой системы у конкретного вида. По соотношению бугорков четверохолмия можно судить о значении для животного слуха и зрения. У копытных и хищных наиболее развиты передние бугорки четверохолмия, которые связаны со зрением, а у китообразных, рукокрылых и утконоса преобладают задние бугорки. Однако крыша среднего мозга у зверей занимает второстепенное положение в обработке как зрительной, так и слуховой информации. Роль ведущих центров начинают выполнять специализированные поля новой коры.
Задний мозг млекопитающих хорошо развит и имеет крупный мозжечок. В отличие от рептилий мозжечок млекопитающих состоит из трех частей: центральной части, или червя, и двух боковых полушарий. У сумчатых, грызунов и насекомоядных размер червя не уступает каждому из полушарий, тогда как у приматов и хищных по размерам доминируют полушария мозжечка. Нужно подчеркнуть, что мозжечок не является ни сенсорным, ни моторным образованием. Он стоит в стороне от главных входных и выходных путей и служит структурой, координирующей двигательную активность с нервной деятельностью других отделов головного мозга.
Внутри головного мозга находится сложная система мозговых желудочков. Полость спинного мозга переходит в ромбовидную ямку продолговатого мозга, которая носит название IV мозгового желудочка. Его передний край прикрыт у большинства млекопитающих мозжечком. Кпереди от мозжечка IV желудочек переходит в сильвиев водопровод — канал, проходящий под средним мозгом и ведущий в III желудочек. III желудочек представляет собой довольно узкое пространство между внутренними стенками промежуточного мозга. Спереди через монроевы отверстия он сообщается с двумя боковыми желудочками. I и II желудочки лежат внутри полушарий переднего мозга. Впереди III мозгового желудочка вследствие местного срастания полушарий вокруг треугольного пространства образуется так называемый пятый мозговой желудочек. Этот мозговой желудочек свойствен только млекопитающим.
Черепно-мозговые нервы представлены 12 парами. Обонятельный (I) нерв обычно выходит из более или менее крупной обонятельной луковицы в виде отдельных небольших волокон.
Только у утконоса он представляет целый ствол, а у дельфинов и китов отсутствует. Зрительные нервы (II) образуют сложный перекрест, а тройничный нерв (V) обычно выходит из черепа отдельными ветвями, которые различаются по функциям. Лицевой нерв (VII) млекопитающих сильно развит, так как является двигательным нервом мимической мускулатуры. Блуждающий, языкоглоточный и добавочный нервы (IX—XI) выходят из черепа совместно. Блуждающий нерв, как и у птиц, простирается далеко в туловище, иннервируя внутренние органы.
Симпатическая нервная система хорошо развита и образует много сплетений. Самое крупное из них находится в передней части брюшной полости позади желудка. Большой симпатический нерв образует парную цепь ганглиев по обеим сторонам позвоночника, которая продолжается до области тройничного нерва и формирует ганглии для его первой, второй и третьей ветвей. В шейной области ганглии немногочисленны, нервы формируют редкую сеть.
Органы чувств у млекопитающих более развиты, чем у всех остальных позвоночных. Для осязания служит кожа, богатая свободными нервными окончаниями и специализированными рецепторами тепла, холода, вибрации и давления. У млекопитающих хорошо развит орган обоняния. В большинстве случаев он состоит их двух частей: обонятельного эпителия, приспособленного для восприятия большинства запахов, и якобсонова органа, который используется для рецепции половых запахов. Слуховая система развита благодаря усложнению строения улитки и развитию коры. У млекопитающих улитка впервые закручивается в спираль, которая у китообразных делает 1,5 оборота, у человека — 2,5, у кошки — 3, у свиньи — 4, а у некоторых грызунов — даже 5 оборотов. Способностью воспринимать электрические сигналы обладают водные позвоночные животные. Ответы электрорецепторов клюва утконоса были обнаружены при изменении напряженности электрического поля. Утконос может воспринимать не только напряженность, но и частотные характеристики электромагнитного поля. Эти особенности позволяют ему находить пищу в непрозрачной воде. Зрение млекопитающих позволяет им различать цвет и форму объектов намного лучше, чем животным других классов. Это достигается как сложностью строения глаза, так и наличием корковой части зрительного анализатора. Цветовым зрением обладает большая часть млекопитающих, хотя у видов, ведущих сумеречный образ жизни, цветовое восприятие понижено.
Сложность поведения млекопитающих не нуждается в подробном иллюстрировании. Большинство млекопитающих могут прогнозировать результат своих действий, выражать эмоции, играть, устанавливать иерархические отношения в группе, обучаться и решать сложные экспериментальные задачи. Коллективные действия волков, крыс и копытных, забота о потомстве, межвидовой альтруизм и заметные индивидуальные особенности поведения конкретных особей приближают млекопитающих к человеку. Морфологические различия между человеком и высшими млекопитающими не носят принципиального характера. Мозг писателя А. Франса весил 1020 г, а мозг средней гориллы — 940 г. Извилин у дельфинов примерно вдвое больше, чем у человека, при равной массе тела. Толщина коры ряда грызунов намного больше, чем у человека и обезьян, хотя мозг полностью лишен извилин. Тем не менее только человеческий мозг способен создавать то, чего до него не было в природе.
