
BIOLOGIA AXIOMATICA
АКСИОМЫ БИОЛОГИИ
Из второго начала термодинамики, о котором говорилось в начале этих очерков, вытекает чрезвычайно важное следствие: нельзя получить энергию только отнимая у какого-либо тела тепло. При этом нужно нагревать какое-то другое тело.
Помню, как вблизи экватора мы поднимали с километровой глубины большой батометр: из него потекла ледяная вода, а на поверхности температура была за тридцать. После вахты я на досуге стал изобретать некий генератор энергии. Состоять он должен был, насколько помню, из двух радиаторов, соединенных двумя трубами километровой длины. Легкокипящая жидкость, наподобие фреона, вскипала в верхнем, плавающем на поверхности моря, радиаторе; пары ее, прокрутив по дороге турбину, конденсировались в нижнем. Такой генератор охлаждал бы поверхностную воду и прогревал глубинную. Не будь солнца, он, конечно, остановился бы, как только температуры наверху и внизу сравнялись. Система пришла бы в равновесное, самое вероятное состояние. Теплота самопроизвольно не может переходить от холодного тела к менее холодному. Наоборот — пожалуйста.
Но теплота — это кинетическая энергия хаотически движущихся молекул. Система «паровой котел — холодильник» с разницей температур менее вероятна, чем система, во всех частях которой средняя кинетическая энергия слагающих ее молекул одинакова. Во втором случае максимума достигает энтропия — так называют омертвленную энергию, которую нельзя превратить в работу. А энтропию физики определяют как логарифм вероятности состояния системы. Хаос вероятнее структуры — люди понимали это всегда. Именно ясным сознанием того, что порядок не может возникнуть из беспорядка, объясняется столь раннее становление двух противоборствующих теорий развития — преформизма и эпигенеза (смотри аксиому первую). Но, пожалуй, лишь великий физик Людвиг Больцман в прошлом веке выразил это количественно.
И невозможность создания вечного двигателя второго рода, при работе которого возникает, а не выравнивается разность температур (или давлений, или электрических потенциалов), стала уже следствием более общего закона самопроизвольного перехода порядка в беспорядок.
Все мы знаем, что для наведения порядка в квартире или хотя бы на письменном столе требуется затратить какую-то долю энергии. А вот беспорядок возникает сам, его специально создавать не нужно.
Чтобы построить жилище — от первобытной хижины до высотного дома, — люди затрачивали уйму энергии. А судьба их творений, если их предоставить самим себе (то есть не тратить энергии на поддержание структуры), была одинаковой: они рассыпались, превращаясь в бесформенные, но зато более вероятные с точки зрения термодинамики груды строительных материалов. Энергия, затраченная на их сооружение, в конце концов переходила в энергию теплового движения молекул, омертвлялась.
А сами структуры наших тел? Не начинают ли они в среднем после 50 лет постепенно распадаться, уступая непреложному росту энтропии? В конечном счете все успехи геронтологии лишь чуть-чуть затормаживают этот процесс. Об этом хорошо сказал Омар Хайам:
| В этом мире ты мудрым слывешь? |
| Ну и что? |
| Всем пример и совет подаешь? |
| Ну и что? |
| До ста лет ты намерен прожить? |
| Допускаю. |
| Может быть, до двухсот проживешь. |
| Ну и что? |
Есть, по меньшей мере, добрая сотня гипотез (по некоторым подсчетам, их более двухсот) объясняющих, что такое старость и как с нею бороться. Я с трудом удерживаюсь от желания добавить к ним сто первую, которая кажется мне наиболее обоснованной. Но сейчас не это является нашей задачей. Мы должны понять, что второе начало термодинамики требует постепенного разрушения генетических программ наших клеток. Случайные, непредсказуемые и равнодушные к судьбам организмов изменения генетических программ приводят к нарушению стройного порядка поддержания фенотипов. «Жить значит умирать» (Ф. Энгельс).
Но этого мало. Ведь клетки — предшественники гамет (яйцеклеток и спермиев) — также подвержены мутациям, которые, буде это случится, перейдут в следующее поколение. Дети передадут их внукам добавлением новых (вспоминаются древнеримские стихи: «Отцы были хуже, чем деды — нас негодных вырастили»). Получается, что игра в испорченный телефон от поколения к поколению в конце концов приведет к полному разрушению структур наших организмов, торжеству энтропии.
Итак, мы пришли как будто к печальному выводу: хаос побеждает структуру. Случайные, ненаправленные изменения генетических программ должны, накапливаясь из поколения в поколение, разрушать и сами программы, и те фенотипы, которые этими программами кодируются.
Если бы вывод был только печален. Но ведь он и неверен! Все мы отлично знаем, что структуры живых организмов сохраняются от поколения к поколению. Более того, мы знаем, что в ходе эволюции шло непрерывное усложнение структур. Два миллиарда лет назад на Земле обитали лишь бактерии и синезеленые водоросли. Каких-нибудь 200—300 миллионов лет спустя появляются организмы с оформленным ядром, пока еще простейшие одноклеточные, примитивные грибки. А дальше — кишечнополостные, черви, моллюски. 500 миллионов лет назад на сцену жизни выходят хордовые. «Век рыб» сменяется «веком земноводных», затем «веком рептилий» и, наконец, на высшие ступени в биосфере выходят млекопитающие, затем человек. Где же здесь победа энтропии?
Да и подчиняется ли живая природа второму началу?
Чтобы решить эту проблему, вернемся назад. В прошлом веке великий физик Максвелл предложил поставить мысленный эксперимент. Представьте себе, говорил Максвелл, трубку, заполненную разреженным газом. Трубка посредине разделена перегородкой. В перегородке есть заслонка, дверца. Допустим, что у этой заслонки сидит некоторое существо (или устройство), различающее молекулы по скоростям. Пусть это существо (демон) открывает заслонку перед быстрыми молекулами и закрывает перед медленными, то есть сортирует их по энергиям.
В результате отбора, производимого демоном, быстрые молекулы соберутся в одной половине трубки, а медленные в другой. Один конец устройства разогреется, другой охладится. Общая энергия устройства останется прежней, так что первое начало термодинамики (закон сохранения энергии) мы не нарушим. Но система перешла от более вероятного состояния к менее вероятному. Демон получил разность температур, позволяющую совершить работу, в обход второго начала термодинамики.
На рисунке показана схема, позволяющая воссоздать парадокс 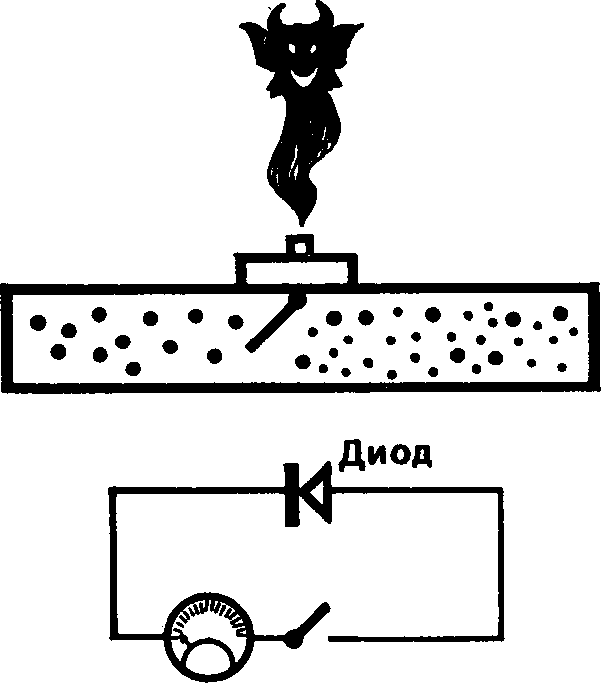 Максвелла. Соберите электрическую цепь с диодом (или иным выпрямителем), пропускающим ток только в одном направлении. Свободные электроны в металле находятся в состоянии хаотического теплового движения (электронный газ). Как и во всяком газе, в нем возникают флуктуации мы их не видим, но слышим как шипение и потрескивание в динамике приемника (там они усиливаются, и они-то как раз являются помехой слабого сигнала). Диод должен сыграть роль демона: пропуская электроны в одну сторону, он создает разность потенциалов, за счет которой можно выполнить работу.
Максвелла. Соберите электрическую цепь с диодом (или иным выпрямителем), пропускающим ток только в одном направлении. Свободные электроны в металле находятся в состоянии хаотического теплового движения (электронный газ). Как и во всяком газе, в нем возникают флуктуации мы их не видим, но слышим как шипение и потрескивание в динамике приемника (там они усиливаются, и они-то как раз являются помехой слабого сигнала). Диод должен сыграть роль демона: пропуская электроны в одну сторону, он создает разность потенциалов, за счет которой можно выполнить работу.
Собрать такую схему в школьном кабинете физики — минутное дело. Еще быстрее мы убедимся, что диод не желает быть демоном — ток в схеме не возникает. Причина? Вернемся к примеру с разреженным газом в трубке. За неимением демона снабдим заслонку какой-нибудь пружиной, которая позволит дверце открываться только после удара быстро движущейся молекулы с высокой энергией. Дверца откроется, чтобы пропустить молекулу, но при этом отнимет у нее энергию на деформацию пружины! В случае с электронами роль пружины выполняет сопротивление диода.
А если у нас все-таки есть демон? Допустим его существование, ведь эксперимент у нас мысленный. Можно допустить даже и то, что на открывание заслонки энергия не расходуется. Но каким образом наш привратник узнает, какую молекулу надо пропустить, а какую нет? Он должен знать скорость молекул, непрерывно получать информацию об их координатах в каждый момент времени. Но информация не дается даром: Л. Бриллюэн показал, что затраты на различение молекул с лихвой компенсируют возможный прирост энергии. Как не вспомнить шутливое замечание о том, что первое начало термодинамики утверждает, что в игре с природой нельзя выиграть, а второе — что нельзя даже остаться «при своих». На атомно-молекулярном уровне отбор оказывается невозможным.
А в живой природе? Тут вступает в действие принцип, который Н. В. Тимофеев назвал принципом усилителя. Правильнее называть его принципом усиления так как под словом «усилитель» обычно подразумевают какое-либо устройство, специально созданное для этой цели. Понять его действие можно из примера, приводимого В. А. Ратнером. Допустим, мы имеем оплодотворенную яйцеклетку — носительницу мутации какого-нибудь гена, кодирующего важный для жизни фермент. В процессе роста и развития организма яйцеклетка превратилась в миллион миллиардов клеток (1015). Соответственно умножились гены. Каждый ген продуцирует, допустим, сто молекул мРНК и на каждой молекуле мРНК синтезируется в среднем сто молекул фермента. Наконец, каждая молекула фермента в минуту осуществляет, скажем, 10 000 актов какой-либо реакции. Итак, 1015•102•102•104 = 1023. Вам, должно быть, известно число Авогадро: количество молекул в моле примерно равно 6 • 1023. Вот насколько усиливаются результаты одного-единственного квантового скачка одной мутации!
Это уже ощутимые количества, с которыми может работать демон. И такой демон существует — это естественный отбор, «демон Дарвина», как удачно его назвал известный биохимик, популяризатор и фантаст Айзек Азимов. Именно отбор пропускает в следующее поколение организмы со структурой, не слишком сильно измененной, или с изменением, дающим повышенные шансы на выживание и дальнейшее размножение. Если преимущество обеспечивается усложнением организации — что же, демон Дарвина отберет и пропустит через свою «заслонку» в будущее чрезвычайно редкие варианты, такие, которые редки, как сверхбыстрые молекулы в газе. Так идет прогрессивная эволюция.
Значит ли это, что жизнь не подчиняется второму началу термодинамики, что она не повышает, а понижает энтропию? Высказались и такие мнения. Но это заблуждение. Жизнь нарушает второе начало не большей мере, чем радиоприемник. Все мы по печальному опыту знаем, что банальный транзистор, приняв невообразимо слабый сигнал, может его усилить до такой степени, что возникнет опасность для барабанных перепонок соседей. Но на это затрачивается свободная энергия батареек. Она расходуется, в частности, на преодоление сопротивления диодов и прочих элементов, превращаясь в тепло — хаотическое движение молекул. Из-за угла снова выглядывает ухмыляющаяся энтропия.
В жизни то же самое. Растения утилизируют лишь немногие проценты падающей на них солнечной энергии (не более двух процентов) Растительноядные животные усваивают не больше 10 процентов энергии пищи, хищники, находящиеся на концах пищевых цепей, — и того меньше. Коэффициент полезного действия жизни существенно меньше КПД первых паровозов Стефенсона. Ничего не поделаешь — на усиление требуется энергия. Жизнь вообще и человек в первую очередь в той же мере снижает энтропию Солнечной системы, в какой карманный воришка повышает национальный доход. Один перераспределяет энтропию, другой — деньги в свою пользу, а в окружающей их среде количество энтропии соответственно возрастает, а денег — уменьшается.
Важно подчеркнуть, что отбор действует не прямо на измененные генетические программы, а на фенотипы, в которых каждое изменение в миллиарды миллиардов раз усиливается.
Вот мы и подошли к формулировке четвертой аксиомы.
Важно подчеркнуть, что естественный отбор, так же как искусственный, где роль условий внешней среды выполняют требования человека,— не просто уничтожение одних особей в популяции и сохранение других. Это дифференциальное размножение, большая вероятность оставить потомство. Вот простой пример: допустим, мы отправили в трудное путешествие через горы и пустыни караван из лошадей, ослов и их помесей — мулов, причем все животные навьючены до предела возможностей. Скорее всего, до цели дойдут лишь мулы, сочетающие выносливость осла и силу лошади. Но они бесплодны, оставить потомство не могут. Это не отбор.
Один весьма уважаемый мною физик эмоционально отрицал значение отбора. Среди его доводов был и такой: уничтожая худшие фенотипы, отбор не улучшает популяцию. Например, у меня в кармане есть золотые, серебряные и медные монеты. Если я выброшу все медные монеты, стану ли я богаче?
Маститый физик не учел одного признака, отличающего монеты от живых организмов. Монеты не размножаются в карманах (в своих я, например, наблюдал только обратный процесс). Но допустим, что они размножаются делением, как бактерии, хотя бы раз в сутки. Если карман имеет неограниченную вместимость (бездонный), то ничего, кроме разрастания нашей «популяции монет» не произойдет. Но бездонных карманов не бывает, как на Земле не бывает бесконечных пространств для расселения видов. Когда карман переполнится, лишние монеты будут высыпаться, рост популяции стабилизируется. Введем фактор отбора выбросим медяки, Освободившееся место займут золотые и серебряные монеты. Выбросим и серебро. Через два-три деления карман будет набит золотом: все-таки мы стали богаче, хотя бы и в мыслимом эксперименте. Еще и еще раз подчеркнем: отбор не уничтожение, а дифференциальное размножение!
Возражение моего оппонента из тех, что юристы называют argumentum ad hominem (апелляция к чувствам). Ведь каждому из нас жалко выбросить хотя бы пятак.
Есть и более сложные, богато снабженные формулами возражения. Берется какая-нибудь математическая модель. В нее подставляют обычно взятые с потолка коэффициенты отбора (так как все попытки экспериментально их определить можно пересчитать по пальцам), производят соответствующие расчеты — и победно заключают, что отбор не может обеспечить эволюцию, демон Дарвина чересчур медлителен.
Что можно сказать но поводу этих моделей? Дифференциальные уравнения в них верны в отличие от исходных положений. Беда в том, что критики дарвинизма переоценивают значение своих моделей. Нельзя отождествлять модель и действительность, портрет всегда будет беднее оригинала.
Рискуя навлечь на себя гнев ценителей прекрасного, назову Венеру Милосскую моделью человеческого организм а (по чресла и без рук). Если случится непоправимое и человечество исчезнет с лица Земли, инопланетный пришелец, найдя в радиоактивных развалинах Лувра знаменитую статую, может по этой модели воссоздать облик человека. Но если он будет утверждать, что люди состояли из однородной массы и не имели ни сердца, ни кишечника, он совершит ту же ошибку, которую делают современные «модельеры» эволюции.
Как правило, это происходит с теми, которые пытаются описать достаточно сложный биологический процесс системой дифференциальных уравнений. Чем сложнее процесс, тем труднее это сделать корректно. На каком-то уровне то приводит к созданию системы нелинейных уравнений, в принципе нерешаемых однозначно. Приходится упрощать задачу, какие-то переменные величины полагать постоянными и какие-то связи между ними — несущественными. Это опасный путь: идя по нему, приходишь порой к результатам, диаметрально противоположным действительности. Казалось бы, следует признать, что модель неадекватна жизни. Но критики дарвинизма верят в обратное и упорствуют в своем заблуждении. И упорствуют они чаще всего только потому, что не желают признать роль случайного в эволюции. Почему же Энгельсу нисколько не претило признание того или иного явления или процесса случайным?
Пожалуй, лучше всего сказал об этом известный венгерский математик Альфред Реньи, приписав свои мысли Паскалю в блестящей стилизации «Письма о вероятности» (Мир, 1970): «...я наткнулся «Размышления» Марка Аврелия и случайно открыл ту страницу, где он пишет о двух возможностях: либо мир является огромным хаосом, либо в нем царствует порядок закономерность… И хотя я уже много раз читал эти строки, но теперь впервые задумался над тем, а почему, собственно, Марк Аврелий считал, что в мире господствуют либо случайность, либо порядок и закономерность? Почему он думал, что эти две возможности исключают друг друга?.. В мире господствует случай и одновременно действуют порядок и закономерность, которые формируются из массы случайностей, согласно законам случайного».
Любой дарвинист подпишется под этими словами, что называется, обеими руками. В эволюции участвуют и случайный, стохастический мутационный процесс (изменение генетических программ) и упорядоченный процесс отбора фенотипов по соответствию условиям внешней среды. Всегда ли эти процессы идут рука об руку?
Мы уже рассматривали примеры быстрого и эффективного действия отбора в предыдущей главе (микробы и антибиотики, крысы и антикоагулянты). Рассмотрим теперь обратные примеры. Что будет с какой-либо структурой организма, если отбор по ней перестанет действовать?
Можно предполагать, что в генетических программах, кодирующих построение этой структуры, будут накапливаться неконтролируемые мутации. В результате признак станет изменяться самым случайным образом, превратится в рудимент и, наконец, попросту исчезнет.
Вот несколько примеров действия отбора на разных уровнях начиная с молекулярного.
Известный исследователь Сол Спигелман поставил замечательный опыт — он моделировал дарвиновскую эволюцию в пробирке.
Есть довольно простой бактериофаг Qβ (читается: «ку-бета»). Генетической матрицей его является короткая нить РНК (3600 нуклеотидных звеньев). Попадая в бактерию-хозяина, он начинает синтезировать белки, и в первую очередь фермент, катализирующий матричный синтез вирусной РНК. Спигелман выделил достаточное количество этого фермента репликазы. С помощью репликазы можно синтезировать РНК Qβ в пробирке, достаточно снабдить ее исходным материалом — нуклеотидтрифосфатами, звеньями будущей цепи и матрицей — очищенной РНК.
Когда молекулы РНК размножатся в пробирке, часть матриц можно перенести в следующую. И так до бесконечности. Оказалось, если переносы делаются достаточно часто, через 15 минут, то в популяции молекул ведется самый настоящий отбор на скорость репликации. Через 74 переноса РНК синтезировалась в 15 раз быстрее, потому что стала в 7 раз короче. Естественно, такая РНК может существовать лишь в тепличных условиях пробирки, она уже не может заразить бактерию. Так и болонка не может загрызть оленя в отличие от своего прародителя — волка. Ненужные звенья нуклеотидной цепи были потеряны за восемнадцать с половиной часов.
А если вести отбор на удлинение цепи? Это можно сделать, если пробирку, где идет синтез, погружать кусочки нитроцеллюлозы. Чем длиннее молекула РНК, тем более прочно она сорбируется. Нитроцеллюлозу с прилипшими молекулами переносим в другую пробирку — и, в конце концов, получим удлинение РНК. Этого мало: путем отбора можно получить РНК, устойчивую к антибиотикам, обычно блокирующим ее синтез.
Так же, как РНК фага Qβ теряет ненужные участки последовательностей, многие организмы теряют ненужные им детали своих структур. Теряют глаза пещерные, подземные и глубоководные животные; потеряли гемоглобин антарктические белокровные рыбы (в холодной воде газы растворяются лучше, а обмен веществ заторможен). И дело здесь не в упражнении органов, как говорят сторонники Ламарка. Отбор перестает следить за формированием структуры, и этого достаточно. Остальное делает второе начало термодинамики.
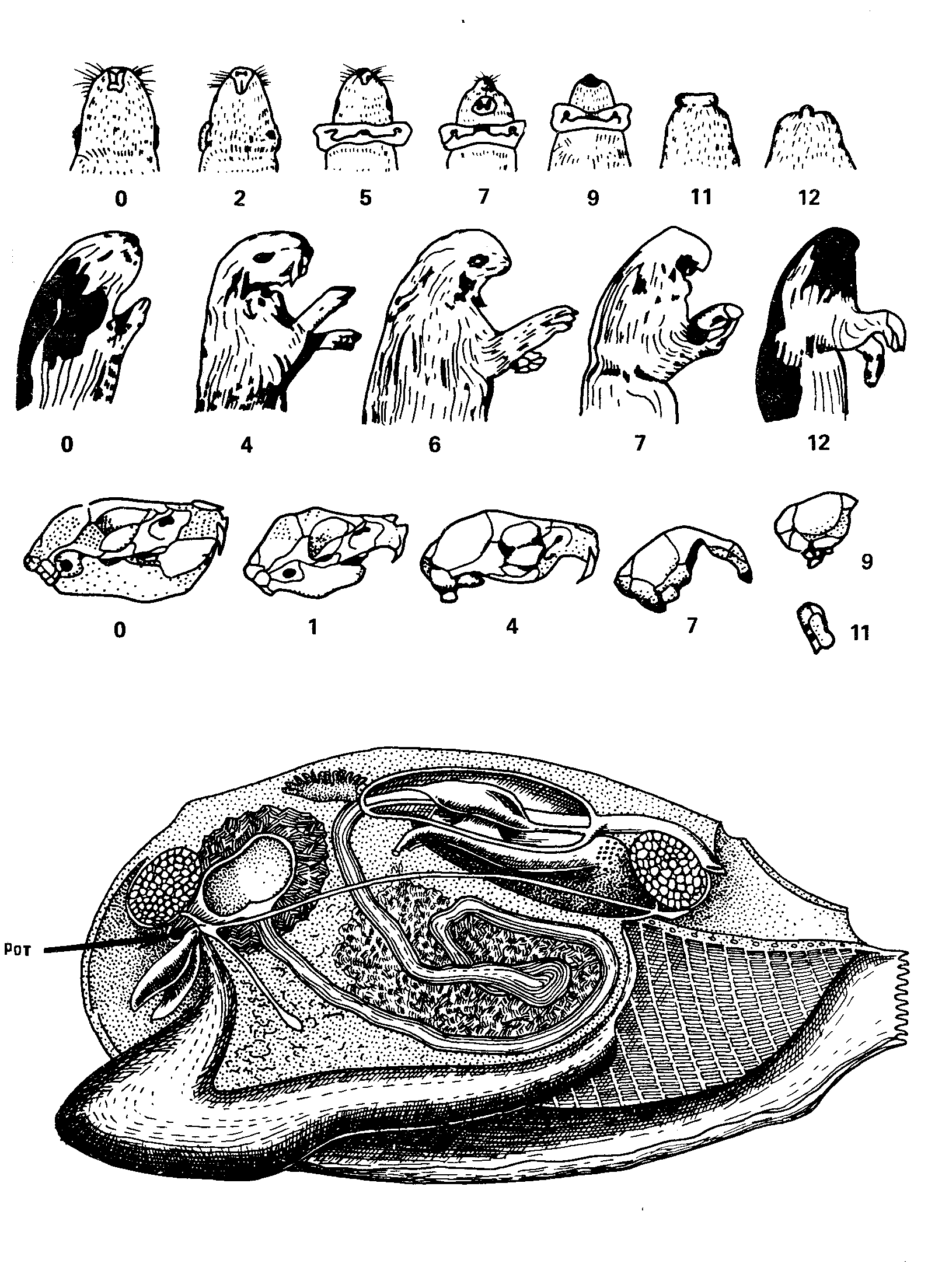
Рис. 38. Вот пример, когда отбор не мешает второму началу термодинамики. У морских свинок имеется серия мутаций, приводящая к частичной (1—4) и почти полной редукции черепа и всего головного отдела. Естественно, у морских свинок такие мутации летальны. Животные с такими дефектами нежизнеспособны. А вот у двустворчатых моллюсков нацело отсутствует голова, и это их не беспокоит (внизу: разрез через беззубку; кишечник, начиная с ротового отверстия, заштрихован). В самом деле, зачем беззубке или устрице голова? Потеря головы также не принесла моллюскам особой пользы (разве что они не страдают мигренями). Но отбор, не следил за ее сохранностью, и постепенно накапливающиеся мутации привели к полной редукции головного отдела. 
Рис. 39. У орхидеи Офрис цветы похожи на самок одиночных пчел и шмелей и пахнут так же. Самцы этих насекомых садятся на цветы, пытаясь с ними спариться, и в результате опыляют. Естественно, непохожие на самок цветы опыляться не будут и потомства не оставят. Но вот один из видов Офриса в Англии превратился в самоопылителя, отбор перестал следить за формой цветов, и они оказались разнообразными по окраске и форме. |
Внутренние паразиты теряют органы чувств, в первую очередь глаза, затем у них до предела упрощается нервная система, наконец, как у ленточных червей, исчезает и пищеварительная. Зачем солитеру кишечник, когда он может всасывать уже переваренную пищу всей поверхностью тела? Но те структуры, которые поддерживает отбор, сохраняются и развиваются, например органы прикрепления к стенке кишечника — присоски, крючья и прочие хитроумные приспособления.
Любопытно, что у личиночных стадий, если они обитают на свободе, такие органы возникают, чтобы потом исчезнуть. У паразита крабов саккулины из яйца выходит свободноплавающая личинка — науплиус (она так похожа на личинок усоногих раков, что сразу выдает происхождение саккулины).
У науплиуса есть глаз, конечности — все, что нужно личинке ракообразного. А взрослая саккулина не похожа не только на свою личинку, но и вообще на рака — это бесформенный мешок, набитый половыми продуктами и прорастающий в ткани краба (сак, саккус — мешок).
Итак, эволюция идет не по отбору сложных, а по отбору приспособленных организмов. Иногда полезна более высокая степень организации, а иногда и наоборот. Н. К. Кольцов писал еще в 1933 году, что «огромное значение регрессивных процессов в эволюции не должно удивлять нас, так как это явление вытекает из применения второго закона термодинамики». Действительно, в случае регрессивного развития отбор не мешает возрастанию энтропии.
По какому основному принципу работает демон Дарвина? Биофизик С. Э. Шноль полагает, что выживают матричные молекулы, способные к более быстрому размножению. Так бывает часто, но не всегда. Слова «более быстрое размножение» не следует понимать буквально. Тише едешь — дальше будешь. Иногда предпочтение отдается не быстроте, а надежности размножения. Вирус, ставший «приживальщиком» клетки, не убивает ее своим стремительным размножением, он приноравливает темп своей репликации к скорости деления хозяина. А если бы он ее убивал, вполне были бы возможны случаи, когда вирус уничтожает популяцию, а затем гибнет сам. Это подтверждают неудачи фаговой терапии многих болезней. Фаги, прежде убивавшие дизентерийную бациллу или холерный вибрион, в конце концов переходят на иждивение хозяев, даже становятся им полезными, превращаясь в плазмиды. И случайно ли «командные посты» в биосфере заняли крупные, медленно размножающиеся животные? Слониха носит плод долго, но зато она рождает слона.
Дело в том, что мелкие, быстро размножающиеся организмы более страдают от неизбирательного уничтожения. Разлив реки или лесной пожар уничтожит и лучших и худших представителей мышиного рода, а более быстрый и более сильный олень от такой напасти может и убежать, тогда как медленный и слабый погибнет. Крупные, медленно размножающиеся и медленно растущие организмы проходят наиболее полную апробацию естественным отбором, и это причина их быстрой прогрессивной эволюции. Мелкие быстро приспосабливаются, но сравнительно редко повышают свой уровень организации.
Думаю, рассмотренных примеров достаточно, чтобы показать, каким путем живая природа избегает‚ повышения энтропии и даже понижает ее: ценой повышения энтропии в окружающей среде. Нарушить закон природы можно, только подчиняясь ему.
Принцип селекции шире дарвиновской теории эволюции. На нем основан любой творческий процесс (вплоть до умственной деятельности человека), ведущий к понижению энтропии. В подсознании идет непрерывный и неощущаемый нами самими перебор вариантов, а в результате возникает новая идея. Все другие объяснения этого процесса, право, ничуть не лучше стародавнего «господь надоумил».
И до сих пор справедливы слова Н. К. Кольцова, написанные в 1929 году: «Для нас, верящих в неизменность закона постоянства энергии, термин «творить» может иметь только одно значение: из многих комбинаций выбирать только одну. Поэтому я считаю, что мы и теперь, как 50 лет назад, имеем право спокойно утверждать: «естественный отбор творит новые формы».
Нам предстоит рассмотреть еще два аспекта деятельности демона Дарвина. Непонимание или игнорирование их приводило и приводит к серьезным ошибкам.
Над этим следует подумать. Во всех ли случаях безотказно работает демон Дарвина? Естественно, он бессилен, когда не из чего выбирать, когда нет хоть малого количества отклонившихся от нормы генетических программ. Такие популяции бывают — это чистые линии организмов, полученные при помощи близкородственного скрещивания или же размножаемые вегетативно. Но мутационный процесс поспешит доставить материал, и через несколько десятков поколений линия утратит чистоту.
Гораздо труднее другая проблема. Я уже упоминал о расчете Манфреда Эйгена : количество вариантов молекул цитохрома C настолько велико, что для его реализации не хватит массы Вселенной. А реальный цитохром C отнюдь не столь изменчив. Это небольшой белок, содержащий чуть более сотни аминокислотных остатков. Оказалось, что 16 кодонов гена цитохрома мутируют с высокой скоростью, 65 — со скоростью в 3,2 раза меньшей, а 32 не мутируют совсем. Ясно, что мутации в них все-таки происходят, но тогда белок перестает выполнять свою функцию быть переносчиком электрона в циклах окисления и организм — носитель мутации — погибает от удушья.
Неужели же эта последовательность в 32 аминокислоты — единственная и неповторимая? думать так — значит признавать существование у истоков жизни некоего разумного творца, избравшего из неизмеримого множества случайных вариантов стабильное ядро цитохрома. Ибо отбору на это времени явно не хватит. Ясно, что среди почти бесконечного числа вариантов цитохромов есть столь же бесконечное количество способных выполнять свою функцию, причем, быть может, лучше, чем реализованная природой. Почему же отбор здесь останавливается?
Начнем издалека. Одно время, помню, была популярна такая игра: превращать муху в слона. Берется какое-нибудь слово и путем изменения букв в нем превращается за несколько этапов в другое. Правила просты: нужно каждый раз изменять только одну букву и каждый раз получать слово со смыслом ( существительное в единственном числе и именительном падеже). Например, слово «лист» превращается в «розу» таким путем: лист — лиса — киса — коса — роса — роза.
Игравшие в такую игру помнят, что далеко не все подобные превращения возможны. Порой с досадой убеждаешься, что путь лежит через бессмысленное буквосочетание или же нужно изменить не одну букву за один этап, а больше.
Примерно такие же трудности испытывает и эволюция. Чтобы получить лучший вариант, нужно сначала провести на следующий этап бессмысленную последовательность, а этого отбор не допустит. Приходится оставлять старую, ведь хотя бы она справляется. Выходит, что демон Дарвина только потому не всесилен, что чересчур ретив. Правда, в одном гене могут возникнуть сразу две, а то и более мутаций, но вероятность этого мала. Если, например, вероятность одной мутации в гене 10-5 то двух: 10-5 • 10-5 = 10-10. Такие ничтожные вероятности не столь часто реализуются.
Генетик С. Райт представил этот парадокс в виде очень наглядной картины. Вообразим разные степени приспособленности к внешним условиям в виде холмистого ландшафта, где высота холма (адаптивного пика) соответствует степени приспособления. Популяция, поднявшаяся на маленький пик, не может сменить его на большой, стать более приспособленной, ибо при смене пиков отбор пойдет против уровня приспособленности. Так, кошка во время наводнения, спасаясь на низком заборе, может утонуть, хотя бы рядом был высокий дом. В конечном счете все вымершие группы (панцирные рыбы и динозавры, саблезубые тигры и мамонты) повинны перед эволюцией лишь в том, что выбрали неудачные, невысокие адаптивные пики. Нам, приматам, повезло, однако надо помнить, что и наш пик не бесконечно высок.
Неужели же нет способа обойти выявленный нами запрет, перехитрить чересчур прямолинейного демона? Прежде всего напомним, что условия внешней среды меняются и то, что раньше считалось неадаптивным, бессмысленным, может оказаться приспособительным. Так и в нашей игре в слова — сочетания букв, прежде не имевшие смысла, входят в язык (в детстве я, например, не смог бы использовать в этой игре слова «радар» или «лазер»). Вспомним о том снижающем жизнеспособность гене, который оказался причиной появления суперкрыс как только вошли в практику антикоагулянты. А вот еще более наглядный пример. Белок, слагающий сокращающиеся нити, которые растягивают хромосомы по разным половинам делящейся клетки, чудовищно консервативен. В основных своих чертах он един для всех ядерных организмов (значит, возник не позже 1,8 миллиарда лет назад). Об этом можно судить по тому, что на клетки всех животных и растений действует колхицин — вещество, выделяемое из лилейного растения безвременника. Колхицин блокирует сокращение нитей, и все хромосомы остаются в одной клетке (на этом основан один из методов получения полиплоидов). Но чтобы получить такой эффект на клетках сирийского (или золотистого) хомячка, требуется концентрация колхицина на два порядка большая. А дело в том, что сирийский хомячок сформировался как вид в бесплодной пустыне, где единственной пищей для него порой был безвременник.
Но во всех этих примерах решающее слово имеет отбор. Есть ли такие ситуации, где демон Дарвина закрывает глаза? Вспомним, что у высших организмов клетки имеют двойной набор хромосом, двойной набор генов. Они диплоидны. Значит, один ген из двух может измениться в неприспособительную сторону и отбор этого не заметит. А в дальнейшем, при изменении условий, это изменение может и пригодиться.
Я уже напоминал, что высшие растения имеют в своем цикле два поколения : спорофитное (диплоидное) и гаметофитное (гаплоидное, с одним набором генов) У подавляющего большинства их гаметофит редуцирован, а то и паразитирует на спорофите, став частью цветка. Все превратности судьбы испытывает диплоидный спорофит А вот мхи — растения наоборот. У них спорофит развивается на гаплоидном гаметофите, каждая мутация сразу идет на апробацию естественным отбором. Вот всем известный мох — кукушкин лен. У него на верхушке женского гаметофита вырастает спорофит, косо наклоненная коробочка на длинной ножке, получающая питание от гаметофита.
И хотя мхи, особенно сфагновые, чрезвычайно распространились по большим территориям со специфическими условиями (болота, лесная подстилка и т. д.), особых тенденций к прогрессивной эволюции у них не наблюдается. Демон Дарвина отбирает среди гаметофитов особей с одним набором генов. Бактерии и синезеленые водоросли* тоже ведь гаплоидны. Образно выражаясь, у них нет резерва генетической памяти, они не помнят прошлого и не готовятся к будущему (разумеется, это выражение не надо понимать буквально). Потому они и остались теми же, какими были и в докембрии, когда они создавали железные руды Курской магнитной аномалии. Все, о чем мы говорили ранее, можно выразить иначе: будущее в эволюционной судьбе какой-либо группы в значительной мере определяется ее прошлым, то есть унаследованной от предков структурой.
✰ Синезеленые, строго говоря, не водоросли. Это одна из ветвей бактерий. Лучше назвать их цианофитами. Но старое название еще широко распространено, особенно в популярных изданиях.
А нет ли способа как-нибудь избежать этого принципа, сходного с библейским («за грехи родителей потомки отвечают»)? Оказывается, такой способ есть и называется он неотенией.
При неотении не развивается окончательная, дефинитивная структура фенотипа. Способность к размножению приобретает личиночная форма, а в дальнейшем она может развиваться по совершенно новому пути.
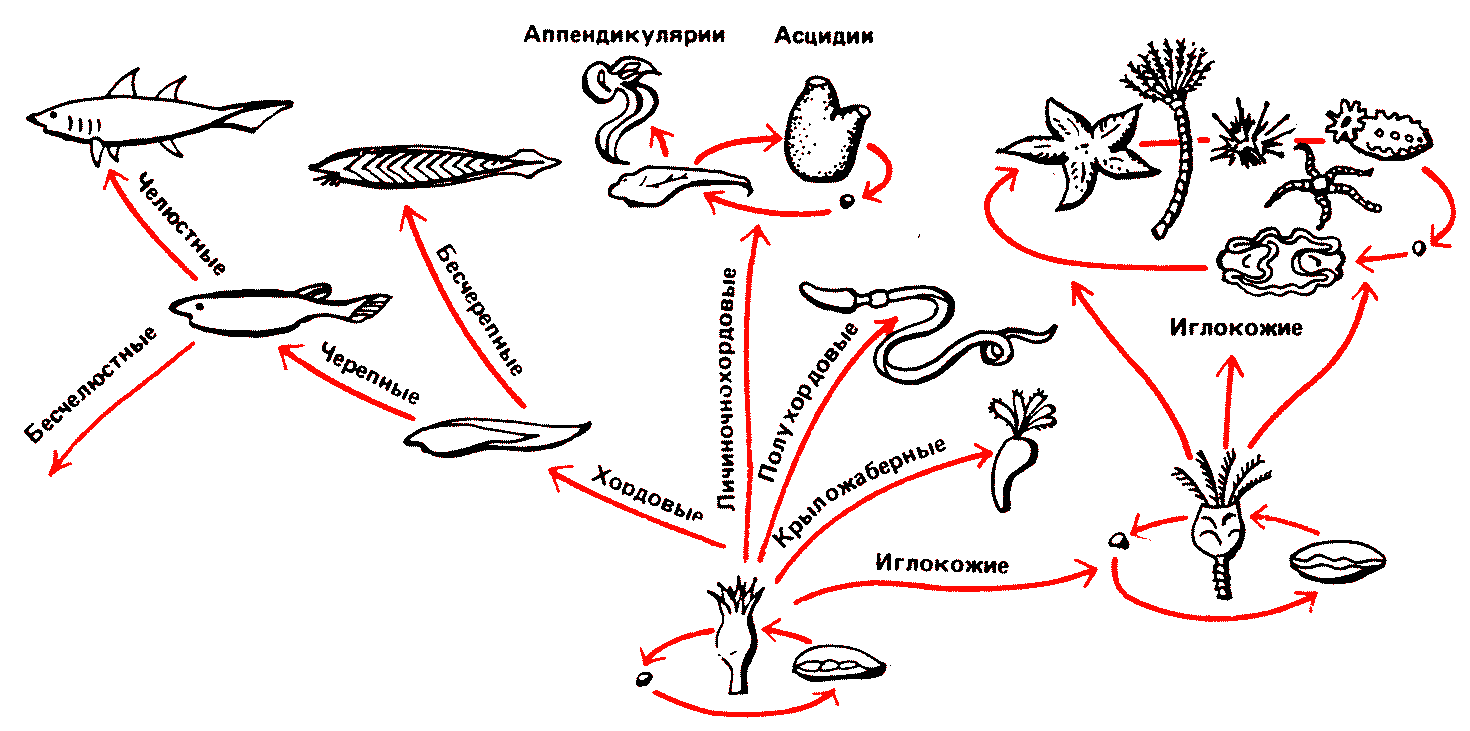
Рис. 40. Считается, что общим предком иглокожих и хордовых был сидячий илоед докембрийской эпохи, радиально симметричный, но имевший в своем развитии стадию двухстороннесимметричной свободноплавающей личинки. Иглокожие — морские лилии, морские звезды, морские ежи, голотурии - так и не сошли с этого пути. И хотя многие из них стали подвижными, они все-таки оказались эволюционным тупиком. Также ничего путного не получилось из крыложаберных, полухордовых и личиночнохордовых. Лишь близкие к асцидиям аппендикудярии потеряли стадию взрослого сидячего организма и стали размножаться, сохранив черты свободноплавающей личинки. Но наибольший успех выпал на долю хордовых. Их размножающаяся личинка в общих чертах, по-видимому, напоминала ныне живущего ланцетника. Затем они обрели хрящевой череп для защиты переднего конца нервной системы, будущего мозга. Но лишь когда передняя пара жаберных дуг превратилась в орган для активного захвата пищи — челюсти, начался триумфальный марш челюстных хордовых: от примитивных хрящевых рыб до нас с вами. |
Быть может, неотения произошла в начале нашего пути — от примитивных животных к человеку. Есть хорошо разработанная гипотеза о том, что хордовые произошли от сидячих илоедов докембрийской эпохи. В их развитии была фаза свободноплавающей личинки. На этом уровне находятся до сих пор живущие в морях оболочники асцидии. У некоторых из них выпала из жизненного цикла стадия сидячего, взрослого организма, личинки стали размножаться сами (на этой стадии находятся также благоденствующие в морях аппендикулярии, они очень похожи на личинок асцидий). Из них-то и возникли примитивные хордовые (и здесь мы можем найти ныне живущую аналогию — всем известного ланцетника). Отсюда уже прямой путь к рыбообразным существам (пока еще без челюстей) эпохи конца кембрия и ордовика. Дальнейшая эволюция вплоть до человека удовлетворительно восстанавливается по ископаемым остаткам. А вот иглокожие, тоже наши родственники, избрали гораздо менее перспективный путь. Здесь демон Дарвина попытался создать из взрослого сидячего илоеда (вроде морской лилии) животное, способное к передвижению. Нельзя сказать, чтобы эта попытка не удалась. Морские звезды, морские ежи и голотурии дошли до наших дней и вымирать не собираются. Но здесь эволюция попыталась создать новую линию на базе взрослой формы, с радиальной симметрией, хотя личинки иглокожих построены по типу более близкой нам билатеральной симметрии (пример радиальной симметрии — снежинка, билатеральной — стрела).
Для сидячего организма радиальная симметрия удобнее, недаром она так широко распространена в мире растений. Но при переходе к подвижному образу жизни она оказалась сущей обузой, и иглокожие, в конце концов, стали эволюционным тупиком. Попытки избавиться от нее на основе плана строения взрослого организма привели лишь к появлению на свет странной формы голотурий и «неправильных» морских ежей. Нельзя вливать новое вино в старые мехи.
Итак, неотения была в начале пути, который вел к человеку. Но она была и в конце его. Человек больше похож на плод человекообразной обезьяны, чем на взрослую гориллу или шимпанзе. Известный антрополог Больк выразил это несколько парадоксально: человек — это половозрелый зародыш обезьяны*.
✰ Впрочем, не все антрололаги разделяют его взгляд.

Рис. 41. Человек — половозрелый зародыш обезьяны. Это звучит парадоксом. Но сравните черепа взрослой гориллы (слева) и ее детеныша. Не правда ли, череп молодой гориллы больше похож на человеческий? Внизу головы зародышей человека (1), гориллы (2) и шимпанзе (З). |
Есть еще несколько способов, так называемых модусов эволюции, которые придают принципу отбора удивительную гибкость, позволяющую создавать новые формы. Но Мы не будем останавливаться на них, так как это, образно выражаясь, теоремы биологии, а не аксиомы ее. Перейдем к одному аспекту четвертой аксиомы, ограничивающему возможности человека, а потому для многих одиозному.
Нам с вами уже трудно представить, какое впечатление произвела на современников ньютоновская теория всемирного тяготения. В то время все казалось возможным, вплоть до предсказания будущего Вселенной. В самом деле, если мы можем рассчитывать движения планет, почему бы не допустить, хотя бы мысленно, такую ситуацию: некий универсальный ум знает координаты и скорости всех тел во Вселенной. Значит, он может рассчитать их местоположение и скорости на любой момент будущего! Это предельный вывод мировоззрения, называемого механическим детерминизмом.
При этом забывали, что даже решение задачи о взаимном притяжении трех тел наталкивается на чудовищные трудности. Находись наша Земля в системе двойной звезды, ее орбита была бы настолько запутанной, что Ньютон просто не смог бы открыть своего закона. Так что притязания механического детерминизма на всеведение оказываются несостоятельными. Тем не менее на макроуровне все процессы детерминированы, и законы Ньютона хотя бы принципиально позволяют предсказывать будущее с предельной точностью (иное дело, что практически это бывает невозможно).
Опустимся на уровень ниже — к молекулам и атомам. Здесь математическое описание совокупности молекул хотя бы в объеме наперстка невозможно. Однако именно из-за большого числа элементов системы предсказание ее поведения возможно. Вместо того чтобы следить за изменяющимися случайным и непредсказуемым образом координатами и энергией каждой отдельной молекулы, мы можем оценить среднее значение параметров системы — температуры и давления. И оценить с высокой точностью: ведь флуктуации, отклонения от среднего значения обратно пропорциональны корню квадратному из среднего. Если в объеме газа содержится 1020 молекул, относительная ошибка будет равна одной десятимиллиардной (10–10). Так что в принципе можно рассчитывать энергию и энтропию системы и предсказывать ее будущее с весьма высокой точностью, как только мы будем оперировать с достаточно большим количеством молекул. Но опять же только в принципе.
Вот пример, близкий каждому. Прогноз погоды, строго говоря, типичная термодинамическая задача. Не приходится говорить о важности прогнозирования погоды и огромных усилиях, которые вкладываются в него. А вот всегда ли эти прогнозы сбываются? Причина та же: невозможно практически учесть все факторы и провести расчеты в обозримые сроки. Детерминизм опять оказывается ограниченным случайностью, это так называемый стохастический детерминизм.
И совсем другое положение в попытках прогнозировать развитие живой природы. В объеме газа случайные флуктуации компенсируют друг друга, на первый план выступает среднее значение. В популяции живых организмов случайное, непредсказуемое изменение генетической программы, усиливаясь в миллиарды миллиардов раз, изменяет состояние всей системы. Вспомните о принципе усиления! Еще один эффектный его пример. В начале 1918 года где-то во Франции квант ультрафиолетового излучения попал на взвешенную в воздухе вирусную частицу — вирион гриппа. Так, а может быть, и иначе, произошла мутация, возник новый штамм вируса, против которого были бессильны защитные системы человеческого организма. Вспыхнула эпидемия, быстро перекинувшаяся на Испанию, оттуда, под названием «испанки» — на весь мир. Смертность достигала 30 процентов, и в результате всемирная эпидемия (пандемия) унесла 20 миллионов человек — много больше, чем первая мировая война.
В пандемии гриппа 1972—1973 годов смертность, к счастью, была не так велика. Но переболело на Земле в то время около двух с половиной миллиардов людей. Нарушение многих планов, отрицательное воздействие на экономику всей планеты, и все это — следствие одного-единственного квантового скачка.
Вывод прост: из-за усиления в фенотипах случайных изменений генетических программ эволюция живой природы принципиально непредсказуема. Наивный детерминизм здесь терпит полный крах. И все, что мы можем предсказать,— это то, что демон Дарвина размножит в будущих поколениях потомков тех особей, которые наилучшим образом будут приспособлены к окружающим их условиям А вот как они будут приспособлены, об этом мы можем только гадать.
Далеко не всем это нравится. Приходится слышать и такие высказывания: современная теория эволюции, объединяющая дарвинизм и генетику, несостоятельна, ибо она может все объяснить задним числом, но ничего не может предсказать. Будь создана теория направленной, номогенетической эволюции, эти недостатки были бы устранены. На мой взгляд, это равносильно утверждению: будь открыт вечный двигатель, с топливным кризисом было бы покончено.
Так что, если мы не желаем отлучить биологию от физики, мы должны признать принципиальную непредсказуемость эволюционных процессов. Кое-что можно предугадать, но это будет достаточно тривиальный прогноз.
Внедряя в практику медицины новый антибиотик, мы с достаточной долей уверенности можем прогнозировать, что он тогда-то потеряет эффективность, микроорганизмы, против которых он направлен, когда-нибудь к нему приспособятся, то есть будут переносить его концентрации, прежде абсолютно для них смертельные. Но вот когда и как? Приспособление может возникнуть разными путями. Один из ферментов бактерии окажется способным к гидролизу антибиотика, повысится непроницаемость оболочки бактерии — это нам знать не дано, ибо это равносильно знанию, когда и в каком месте произойдет событие на квантовом уровне.
Или же другая важная проблема — вредные насекомые и инсектициды. Действуя ДДТ на экспериментальную популяцию мух, легко вывести линию, переносящую без ущерба для себя в сто раз большую концентрацию яда. Но в одной линии это произойдет, потому что ДДТ будет обезвреживаться в мушином организме, в другой — повысится непроницаемость хитинового покрова. Можно, наконец, вывести мух с безусловным рефлексом избегания отравленных поверхностей.
Иногда эволюция кажется направленной, жестко детерминированной. Дельфин, вымерший ихтиозавр и акула очень похожи по форме друг на друга и каждый из них — на подводную лодку. Но зададимся вопросом: а какая еще форма будет отобрана демоном Дарвина для быстрого движения в воде? На некоторые задачи, поставленные перед эволюцией, может быть дан только один ответ.
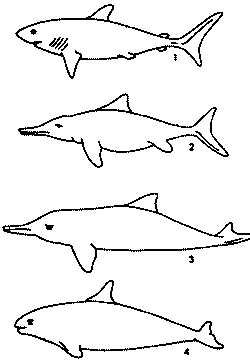
Рис. 46. Жюль-верновский капитан Немо придал своему «Наутилусу» сигарообразную форму, наиболее оптимальную для быстрого движения в воде. Природа использовала этот принцип много раньше.
|
Значит, все-таки результаты эти предсказуемы? Не спешите с ответом: предсказуемость отнюдь не стопроцентная. Вот пример, который можно бы назвать: «Эссе об акульем хвосте». На хвостовом стебле акул (и других быстрых рыб) по бокам расположены кили, отчего в сечении они имеют обтекаемую форму. Такие же кили есть и на хвостовом стебле дельфина, но не по бокам, а сверху и снизу. Понятно почему: у рыб хвостовые лопасти расположены вертикально, а у китов — горизонтально. Этого мало: у акул на нижней стороне хвостового стебля есть поперечная ложбинка. Для чего она, неясно. Тур Хейердал писал, что она для того, чтобы акулу можно было удобнее схватить за хвост. Это, конечно, шутка. Наверное, ложбинка как-то улучшает гидродинамические свойства хвоста. Такие же ложбинки есть и у дельфинов, но по бокам хвостового стебля. Так вот, можно ли предсказать, как расположатся хвостовые лопасти у животного, потомки которого перейдут к быстрому плаванию: вертикально, как у рыб и ихтиозавров или же горизонтально, как у китообразных?
Наконец, у акул последний отдел позвоночника продолжается в верхнюю лопасть хвостового плавника, а у ихтиозавров — в нижнюю.
Смогли бы вы, оказавшись в силурийском периоде, прогнозировать этот результат? Так что даже если ответ один, модификаций его может быть много; столько же, сколько раз решалась в эволюции эта задача.
Иногда же ответов несколько, причем они отнюдь не равноценны. Вот хороший пример. Представьте разреженный лес или саванну, где трава выжигается солнцем. У крупных копытных в этой зоне есть хороший источник питания — листва деревьев. Но как до нее добраться? Самый прямой ответ — отбор на увеличение размеров тела. По этому пути пошли гигантские безрогие носороги олигоцена — индрикотерии. Нельзя сказать, чтобы решение было удачным. Ведь если рост увеличивается, то вес также возрастает пропорционально третьей степени роста. Размеры возросли вдвое, а расходы на питание — в восемь раз.

Рис. 42. Как копытному животному добраться до листьев на высоких деревьях? В процессе эволюции разные животные решали эту проблему по-разному. Самый простой, но не самый выгодный путь избрали гигантские безрогие носороги третичного периода — индрикотерий, белуджитерий и близкие формы. На рисунке реконструкция индрикотерия (Индрик-зверь, царь зверей старорусских преданий) и для масштаба — современный индийский носорог. Такая махина могла легко дотянуться до высоко расположенных веток, но и прокормить ее было трудно. Поэтому гигантские носороги самые крупные наземные млекопитающие — вымерли, не оставив потомства. |
Другие животные вставали на задние ноги, а передними (нередко с мощными загнутыми когтями) подгибали ветки с листьями и плодами. Так вели себя, очевидно, халикотерии, о которых уже упоминалось. Предел этой тенденции — гигантские ленивцы, мегатерии, вымершие в Южной Америке сравнительно недавно, может быть, не без содействия человека.
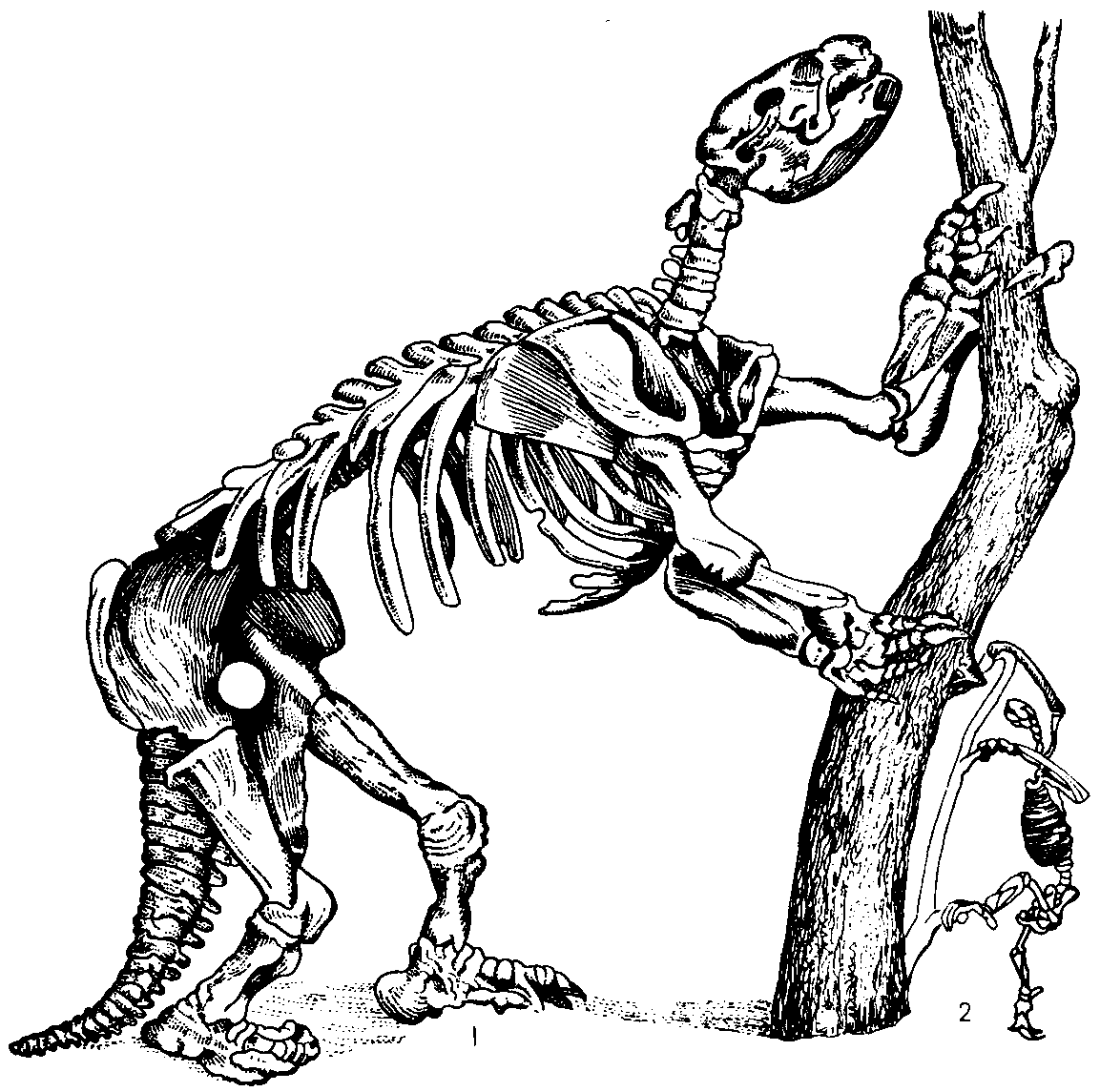
Рис. 45. А вот еще один способ добраться до листьев на деревьях. Гигантские южно-американские ленинцы поднимались на дыбы и подгибали дерево, охватывая ствол передними лапами с мощными когтями. На рисунке скелет ленинца — милодона, раскопанный в Аргентине еще Чарлзом Дарвином, справа (2) скелет современного ленинца. По-видимому, так же вели себя халикотерии, о которых упоминалось выше в связи с ошибкой Кювье. |
У слона появляется длинный хобот, с помощью которого он дотягивается до листвы. Но, по-видимому, наиболее перспективный ответ — удлинение шеи. У всем известной жирафы длинная шея и длинные ноги, что позволяет ей объедать листву на высоте, для других копытных недоступной. К этому еще добавляется длиннейший язык (жирафа и ее родственница окапи могут облизывать себе уши).

Рис. 43. Как мы уже видели, пропорциональное увеличение размеров тела для того, чтобы достать листья с высоких деревьев, энергетически невыгодно. Большего успеха в жизненной борьбе достигли животные со специализированными приспособлениями. Слон дотягивается до веток хоботом (это продукт разрастания носа и верхней губы), у жирафы удлиняются шея, ноги и язык. Геренук (она же жирафовая газель, или антилопа Уэллера) также добирается до листьев на высоких кустарниках с помощью длинной шеи и ног; к тому же она отлично стоит на задних ногах, балансируя передними. |
Этот удачный тип строения был использован эволюцией не один раз. В миоценовом периоде в Северной Америке обитали жирафоподобные верблюды — альтикамелюсы. А в Сомали, в самых жарких и бесплодных местностях, и сейчас благоденствует удивительная жирафовая газель (она же геренук, или антилопа Уэллера). У нее длинная шея и ноги; кроме того, геренук может объедать листья на высоком кустарнике, стоя на задних ногах.

Рис. 44. Внешний вид жирафы известен каждому. Но не каждый знает, что в миоценовом периоде в Северной Америке обитали жирафоподобные верблюды — альтикамелюсы (на рисунке реконструкция внешнего вида). По-видимому, жирафоподобный тип строения оказался очень удачной эволюционной находкой, если повторялся по меньшей мере два раза. И, однако, альтикамеллос остался верблюдом, на жирафу он похож только внешне. |
Вот такие примеры конвергентного, независимого возникновения сходных признаков до сих пор многим кажутся убедительным доказательством направленности эволюционного процесса. Причем направленным считается даже не действие отбора, с чем можно согласиться, а сама изменчивость.
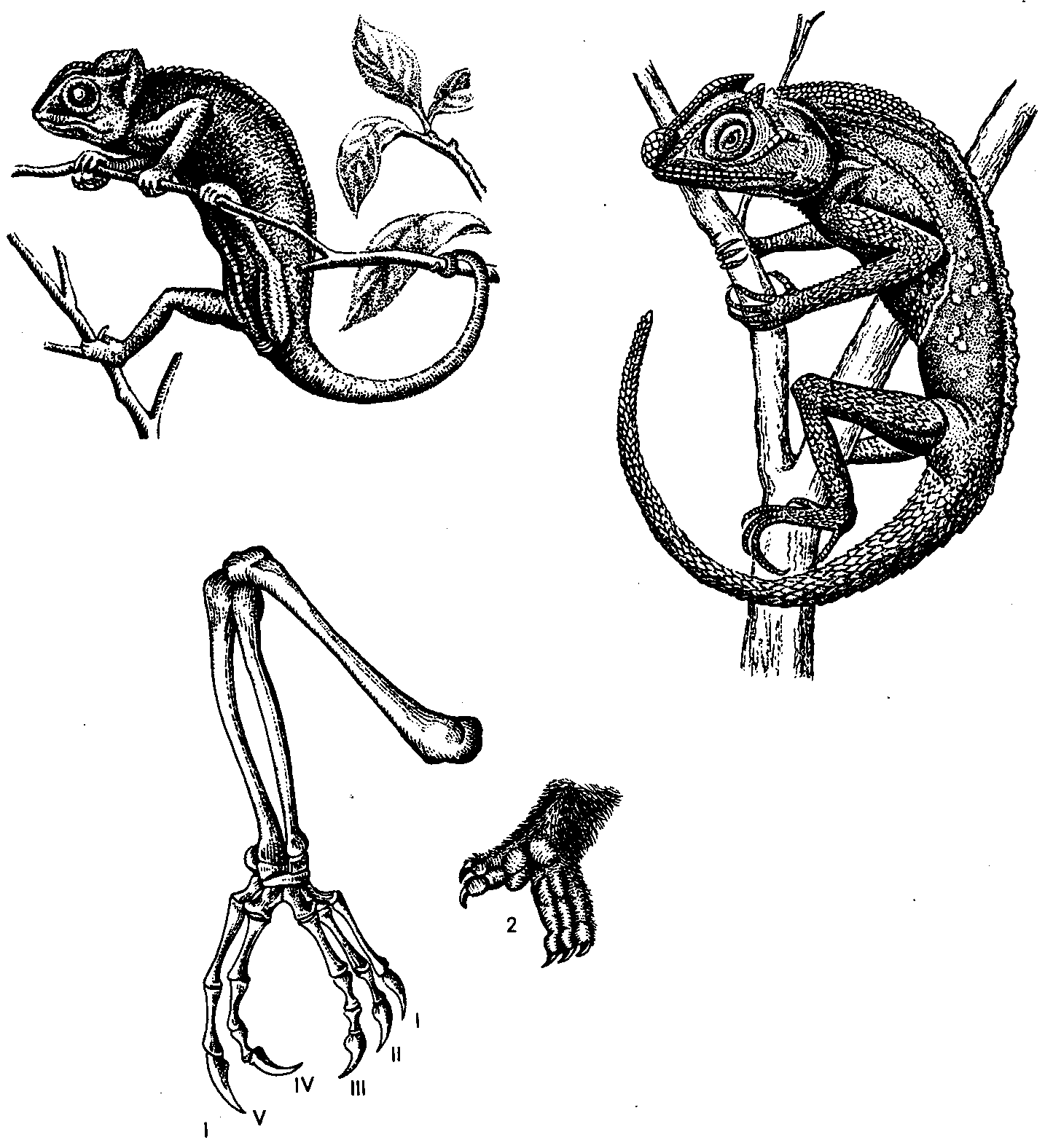
Рис. 48. Животные, обитающие на деревьях, также дают нам яркие примеры конвергенции. Сходный образ жизни приводит к возникновению сходных приспособлений. Слева — хамелеон, справа — цейлонская древолазящая ящерица агама. Их связывает весьма отдаленное родство, но внешний вид сходен. Этого мало: многие детали структуры, например охватывающая ветвь конечность, возникают и у пресмыкающихся и у млекопитающих (внизу скелет конечности хамелеона и лапа кус-куса — древолазящего сумчатого, обитающего на Новой Гвинее). Есть и лягушки с подобными лапами. |
Самые последовательные из сторонников такого толкования полагают, что абсолютно идентичные формы могут возникать из несходных, что один вид может иметь двух, а то и более предков. Это равносильно утверждению, что первоначально несходные последовательности ДНК в результате накопления мутаций станут идентичными и приведут к возникновению идентичных фенотипов. В свете наших сегодняшних знаний о структуре ДНК и механизме ее генетических изменений подобная декларация не заслуживает опровержения. Вероятность того, что два романиста из разных стран независимо друг от друга напишут одинаковые до последней запятой романы, куда выше вероятности конвергентного возникновения видов.

Рис. 50. Еще раз напомним: конвергентными могут быть лишь детали строения организма, но не сами организмы. Здесь изображены тазовые кости динозавра (I), американской птицы тинаму (II) и американского страуса нанду (III). Сходство действительно большое. Но даже самого птицеподобного динозавра струтиомимуса ( само название его — подражающий страусу) нельзя спутать ни с тинаму, ни со страусом. Так же различаются тинаму и нанду. Один и тот же болт может быть использован и в самолете и в автомобиле, но эти структуры легко различаются. |
Еще одно следствие, вытекающее из четырех наших аксиом: конвергентными могут быть только признаки, части структур, но не целые структуры, а тем более не сами организмы.
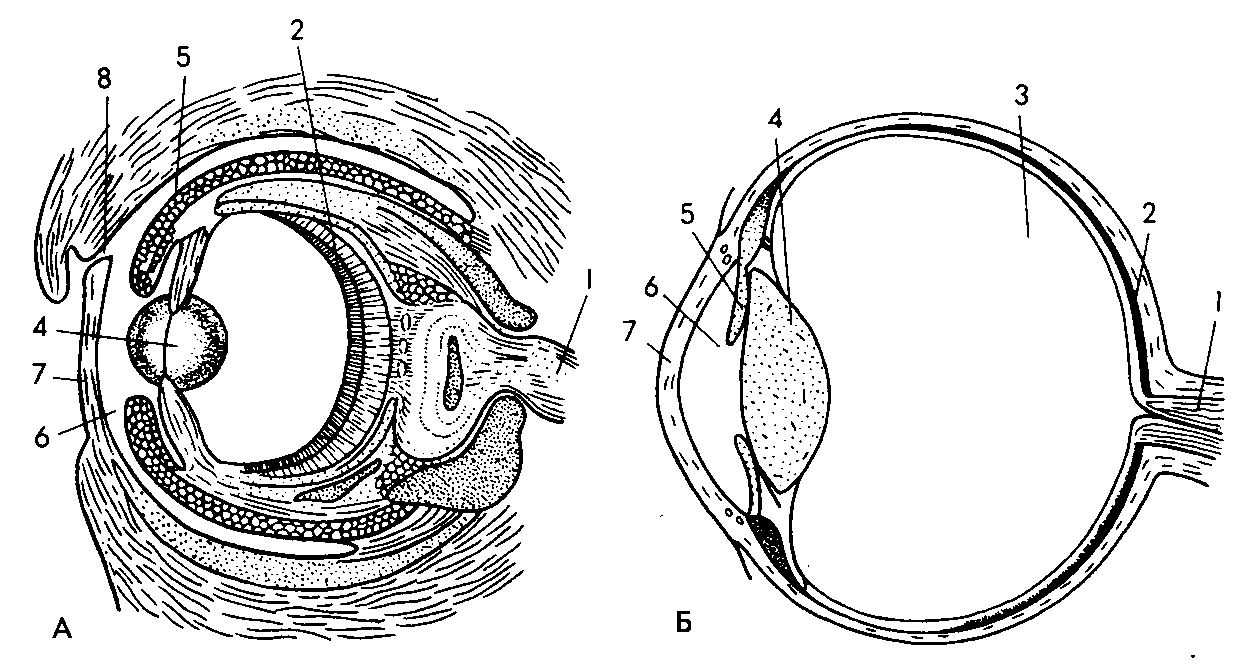
Рис. 47. Один из популярнейших примеров конвергенции — глаз осьминога (А) и глаз человека (Б). Одинаковыми цифрами показаны аналогичные части. Казалось бы, сходство идеальное (у осьминога хрусталик более округлый из-за того, что у воды больший коэффициент преломления, чем у воздуха). Но глаз осьминога «наводится на фокус» приближением или удалением хрусталика от сетчатки, как объектив фотоаппарата. У человека фокусное расстояние изменяется путем изменения кривизны самого хрусталика. Несмотря на внешнее сходство, принципы работы этих оптических приборов совершенно различны. |
Много пишут об удивительном конвергентном сходстве глаза человека и осьминога. Внешне они действительно похожи, вплоть до выражения. Но принципы действия совершенно различны. Глаз осьминога, как объектив фотоаппарата, «наводится на фокус» приближением или удалением хрусталика к сетчатке. У человека фокусное расстояние регулируется изменением кривизны самого хрусталика (представьте оптический прибор с эластичной линзой, фокусное расстояние которой изменяется, например, при накачке в линзу жидкости, сильно преломляющей свет; насколько я знаю, подобный принцип в технике не был использован).

Рис. 49. Конвергенция может сыграть злую шутку с исследователем. Что изображено на этом рисунке? Кроты, скажете вы и будете правы. Только это не наши обычные кроты: вверху изображен златокрот из Южной Африки, тоже насекомоядное, но из другого семейства, а внизу сумчатый австралийский крот. Еще раз подчеркнем: конвергентные формы — это книги разного содержания в одинаковых переплетах. |
Сумчатый крот, обитающий в Австралии, удивительно похож на нашего крота, но у него имеется сумка для вынашивания детенышей и задние концы нижней челюсти, как у всех сумчатых, загнуты внутрь. Это вполне понятно: отбор приводит к конвергентному сходству по признакам, которые полезны в конкретных условиях (например, к подземному образу жизни), но оставляет те, которые могут быть разными без нарушения их функций. Поэтому на Земле нет двух видов с одинаковыми генетическими программами.
Этого мало: можно утверждать, что каждый вид уникален и, раз исчезнув с лица Земли, не возникает вновь. Нельзя воссоздать заново мамонта и Стеллерову корову, среднеазиатского тигра и гепарда, вымерших в последние годы. Даже когда мы научимся сами создавать новые виды, мы можем сделать их лучше вымерших, но идентичных не получим. Потеря каждого вида безвозвратна.
Возможности человека уничтожить любой вид, включая себя, безграничны, возможности исправить содеянное равны нулю. И дело тут не в ограниченности наших знаний, а опять же во втором начале термодинамики, запрещающем воссоздать утерянную генетическую программу.
Обратное положение из этого тезиса: если в геномах двух видов найдется хотя бы одна нуклеотидная последовательность достаточной (более 50 нуклеотидных пар) протяженности, они связаны родством, ведут начало от одного предка. Ибо вероятность возникновения одной последовательности из двух первоначально несходных слишком мала, чтобы ее можно было принимать во внимание. С большей вероятностью вода, выплеснутая из стакана, сама, самопроизвольно соберется в него.
На этом принципе основана новая отрасль систематики — геносистематика, основы которой заложил академик А. Н. Белозерский. И не только геносистематика. Рискну высказать утверждение, что геносистематика, теория эволюции геномов и т. д. в конце концов окажутся лишь частью общей теории эволюции последовательно реплицирующихся систем.
Такой теории еще нет. Однако и сейчас можно полагать, что все системы, передающиеся из поколения в поколение с накоплением ошибок репликации, под контролем внешней среды изменяются, подчиняясь сходным закономерностям.
Под эту категорию попадают не только объекты биологической эволюции, но и, например, человеческие языки, обычаи, обряды, мифы (включая религии), сказки и многое другое. Когда-нибудь я напишу и об этом.



